co ovlivňuje velikost skupin, ve kterých se primáti rozhodnou žít?

Jedním z hlavních teoretických příspěvků studiem primátů je pochopení toho, co ovlivňuje charakter komplexní změnu v sociální struktura a organizace. Základem je soubor teoretického vývoje zkoumajícího, proč zvířata žijí ve skupinách a proč jsou skupiny určité velikosti. Zde se zaměřujeme na otázku „co ovlivňuje velikost skupin, ve kterých se primáti rozhodnou žít?“
primáti jsou zvláště užiteční taxony k řešení této otázky, protože jejich velikosti skupin jsou velmi variabilní, a to nejen uvnitř, ale také mezi druhy. Vnitrodruhových velikost skupiny pro red colobus (Procolobus rufomitratus), například, se pohybuje mezi 12 a 150 členů (Chapman & Chapman 2000a). Mezidruhové variace se pohybují od jednoho jedince, jak je vidět u orangutanů (Pongo sp.) (van Schaik 1999), více než 800 členů v mandril (Mandrillus sphinx) (Aberneth et al. 2002). Kromě toho se v rámci některých druhů sociální skupiny opakovaně rozdělují a znovu se spojují do podskupin různých velikostí a kombinací v průběhu času (např.) a šimpanzi (Pan troglodytes) nebo víceúrovňová organizace gelada (Theropithecus gelada) a paviánů Hamadryas (Papio hamadryas, Aureli et al. 2008). Tato variace poskytuje základ pro výzkumníky k vývoji modelů pro zkoumání jak ekologických, tak sociálních faktorů velikosti skupiny.
seskupování je výhodné několika způsoby. Jednotlivců do větších skupin je myšlenka mít snížené riziko predace (Hamilton 1971), může být lépe schopni najít a bránit potravinové zdroje (Cody 1971, Wrangham 1980), a může být chráněn proti conspecific škodlivý, jako novorozenců pomocí extra-skupina samci (Wrangham 1979). Různí vědci navrhli, že seskupení poskytuje takové předvídatelné výhody (Alexander 1974, van Schaik 1983), že rozdíly ve velikosti skupiny lze vysvětlit nevýhodami (Wrangham et al. 1993). Za nejrozšířenější potenciální náklady na seskupení se považuje snížení účinnosti shánění potravy. S jinými jedinci se stejným dietní požadavky, znamená to, že zvířata buď boj o jídlo (soutěž soutěž), nebo jedno zvíře ve skupině bije další na jídlo, tak při druhé zvíře přichází do oblasti, tam je prostě žádné jídlo vlevo (scramble competition, Janson & van Schaik 1988). V obou těchto situacích se předpokládá, že konkurence o jídlo vede k tomu, že zvířata musí cestovat dále. Logika tohoto argumentu je relativně jednoduchá. Zvířata musí pást na ploše, která splňuje jejich energetické a nutriční požadavky. Z toho vyplývá, že zvýšení velikosti skupiny zvýší oblast, která musí být pokryta, aby bylo možné najít dostatečné zásoby potravin. Jednotlivci tak musí cestovat dále a vynakládat více energie, pokud jsou ve velké skupině, než kdyby se krmili v menší skupině. S nárůstem času stráveného cestováním se blíží bod, kdy energie vynaložená na cestování je příliš nákladná a menší skupiny se stávají výhodnými. Tímto způsobem mohou ekologické faktory ovlivnit pohybové vzorce a efektivitu hledání potravy, čímž omezují velikost skupin, které mohou efektivně využívat dostupné potravinové zdroje. Tyto myšlenky byly formálně v čem stal se známý jako Ekologická Omezení Model (Chapman & Chapman 2000b, Ganas & Robbins 2005, Snaith & Chapman, 2007, Teichroeb & Sicotte 2009).
podstatnou složkou modelu ekologických omezení je, že zvýšení velikosti skupiny musí vést ke zvýšení konkurence v krmivech uvnitř skupiny. Je možné, že to funguje poněkud odlišným způsobem v závislosti na povaze zdrojů používaných konkrétními druhy. S živící se ovocem, a možná mnoho folivorous primátů, které se živí v diskrétní náplasti — typicky stromy nesoucí potravin nebo shluky stromů — další členové skupiny mohou poškozují skvrny rychleji a vést ke zvýšenému den se pohybuje v rozmezí (Chapman 1988, Snaith & Chapman 2005). Pro více hmyzožravé druhy, jejichž zdroje může dojít v jako diskrétní náplasti, neustálé cestování po celém vrchlíku při hledání hmyzu s dalšími členy skupiny může vést k zvýšení překrytí jednotlivých vyhledávacích polí, snížení na obyvatele sazby setkávají s jídlem, a tím se zvyšuje plocha, která musí být prohledány (van Schaik et al. 1983).
MODEL ekologických omezení tedy naznačuje, že ty faktory, které ovlivňují vzdálenost, kterou zvířata cestují, by měly také ovlivnit velikost skupiny. U zvířat, která se obvykle živí ovocem nebo listy, které mohou být vyčerpány, velikost náplasti by určila, jak dlouho může skupina dané velikosti zůstat a krmit. Velká skupina by trávila méně času v náplasti dané velikosti než menší skupina, protože rychleji vyčerpává náplast-velká skupina má jednoduše více úst na krmení. Pokud zvířata cestují mezi náplastmi, jakmile je vyčerpají, hustota a distribuce náplastí určí vzniklé cestovní náklady. Když resource opravy jsou na vysoké hustoty, nebo v denzním distribuce, vzdálenost k další patch je malý, cestovní náklady jsou nízké, a zvířata proto mohou tvořit velké skupiny. V takových časech, jakékoli dodatečné náklady spojené s členstvím ve velké skupině, jako je potřeba navštívit mnoho záplat, lze snadno obnovit. Naproti tomu, když zdroj skvrny se vyskytují v nízkých hustotách, vzdálenost k další patch je obvykle velký, a cestovní náklady jsou vysoké, a zvířata si nemohou dovolit, aby se rychle poškozují záplaty, a proto zvířata tvoří malé skupiny.
fission-fusion sociální organizace pavoučí opice, šimpanzi, a několik dalších primátů (viz Obrázek 1) nabízí užitečné testy ekologická omezení model, protože jeden může pokusit předpovědět časové změny v podskupině velikosti od přímého měření velikosti, hustoty a distribuce potravinových zdrojů (Chapman 1990, Chapman et al. 1995). U pavoučích opic a šimpanzů byl vyvinut vícenásobný regresní model, který ukázal, že velikost, hustota a distribuce potravinových náplastí měřených každý měsíc byly dobrým prediktorem velikosti podskupiny. Je obtížné vizuálně představují vícenásobné regrese, ale metody, které byly použity pro pavouk opice jsou zobrazeny na Obrázku 2. Pavoučí opice mají velmi flexibilní typ štěpení-fúze sociální organizace, a v Národním parku Santa Rosa, Kostarika, velikost podskupiny se může pohybovat od 1 na 35 jednotlivci,ale v průměru o 5 jednotlivci se nacházejí na cestách společně. Pro tuto populaci lze 50% rozptylu průměrné měsíční velikosti podskupiny předpovědět z relativně hrubých opatření velikosti,hustoty a distribuce potravinových náplastí (Chapman 1990).

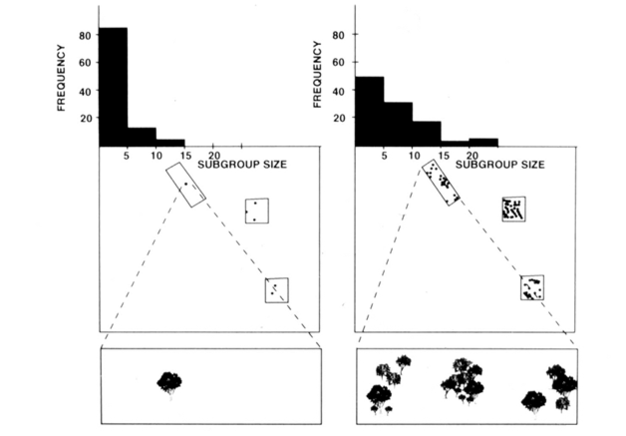
kromě druhů uvedených výše, uplatnění ekologických omezení model byl obecně podporován výzkum z různých druhů (Snaith & Chapman 2005) a situaci (např. obecné modely, Wrangham et al. 1993, and mixed species associations, Chapman & Chapman 2000c). Domníváme se, že vzhledem k rozsáhlé podpoře, kterou tento model získal, nastal čas, kdy je rozumné navrhnout, aby druhy obvykle odpovídaly očekáváním modelu ekologických omezení. To neznamená, že všechny druhy, které budou odpovídat, a tak se stává vzrušující doba pro výzkum v této oblasti, protože vyšetřovatelé mohou hledat výjimky z modelu. Máme podezření, že druhy, které se neshodují, tak učiní, protože přijaly sociální strategie, které jsou v rozporu s ekologickými očekáváními. To poskytuje „měřítko“ pro vyhodnocení potenciálního významu různých sociálních strategií(tj. čím více se odchyluje od očekávání ekologického modelu, tím důležitější jsou sociální selektivní tlaky).
uveďme stručně dva příklady z našeho předchozího výzkumu. Samice šimpanzů byly osamělejší než samice pavoučích opic, přesto mají obě podobné sociální organizace štěpení a fúze. Dále, zatímco my můžeme přesně předpovědět počet mužů a subadult šimpanzi ve skupině na základě ekologických podmínek, jsme byli schopni předpovědět počet samice šimpanzů v těchto podskupinách (Chapman et al. 1995). I když zdroje byly velmi bohaté a téměř všichni muži byli v jedné velké podskupiny, ženy jen zřídka vstoupil skupin, což naznačuje, že náklady jsou ve větší podskupiny převážily jakékoliv výhody, jako je predátor, vyhýbání se daňovým povinnostem. To je neočekávané z pohledu, že kojenci těchto žen by byli věkovou / sexuální třídou nejvíce ohroženou predací. Jedna testovatelná hypotéza k vysvětlení těchto pozorování je, že povaha koalic u těchto dvou druhů může ovlivnit výhody členství ve skupině. Důkazy naznačují, že na rozdíl od šimpanzů vytvářejí samice pavoučích opic koalice, které často fungují, aby umožnily členům koalice exkluzivní přístup k jídlu. Takže pavoučí opice, když se připojí k konkrétním jednotlivcům, mohou zvýšit svůj přístup k jídlu vyloučením ostatních, ale u šimpanzích samic tomu tak není (Chapman et al. 1995).
Jsme identifikovali další zajímavé odchylky, které by mohly být způsobeny sociálními faktory, které ovlivňují náklady a přínosy členství ve skupině, a to se týká red colobus a černá a bílá colobus opice (aka. guerezas, Colobus guereza) v Národním parku Kibale v Ugandě. Červený kolobus tvoří velké skupiny s průměrem 65 jedinců (25-127 jedinců, Snaith et al. 2008), zatímco guerezové žijí v malých skupinách s průměrem 6,5 jedinců (4-11 jedinců, Harris & Chapman 2007). Přes tento rozdíl existuje velká podobnost v rostlinách jedených těmito dvěma druhy. Pokud se ekologické podmínky byly zodpovědné za rozdíl ve velikosti skupiny mezi dvěma colobine druhů, dalo by se očekávat, že jejich strava by se liší a hustota potravin stromy by být nižší v domácnosti rozsahy guerezas, jelikož mají menší velikost skupiny. Jejich strava je však velmi podobná; dietní překrytí mezi červeným kolobusem a skupinou guerezy, která měla svůj domácí rozsah zcela v domácím rozmezí skupiny červených colobus, bylo 43.2%, zatímco pro dvě sousední skupiny red colobus, strava překrývaly pouze 37.3% (Chapman et al. 2002). Zdá se, že žena guereza reprodukční úspěch je maximalizována v malých a středně velkých skupin, a to buď, protože větší skupiny zkušenosti vyšší míru převzetí a novorozenců nebo více krmení soutěže (viz důkaz scramble soutěže v folivorous Trachypithecus phayrei a Semnopithecus sp. , ale nedostatek důkazů o vyčerpání potravinových náplastí guerezas). Jsme svědky počet mužských převzetí a kojenecké vraždy v guerezas (Onderdonk 2000) — viz Teichroeb & Sicotte (2008) pro hodnocení teorie týkající se novorozenců v colobines. Není jasné, jak by riziko infanticidu omezilo velikost skupin guerezy, když mají zjevný přístup k vysoké hustotě potravin, které by měly umožnit tvorbu skupin s více muži. U několika druhů, větší ženské skupiny jsou větší cíl pro emigraci mužů, a tato situace může zvýšit novorozenců sazby-například, Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). Nicméně, zabíjení novorozených dívek se očekává, že budou méně časté, pokud skupina obsahuje více reprodukčně aktivní muže, protože mužské přistěhovalectví a následné násilné převzetí a novorozenců jsou méně pravděpodobné, že dojít, když nováček je konfrontován s několika možných otců budoucích dětí (Janson & van Schaik 2000). Zdá se, že guerezové mají dostatek potravinových zdrojů k vytvoření větších skupin (Tombak et al. 2012), což by snížilo riziko infanticidy, přesto netvoří velké skupiny. Je možné, že povaha interakcí mezi muži a muži v guerezách způsobuje, že velké skupiny jsou nestabilní. To se zdá být případ medvědí colobus (Colobus vellerosus), kde multi-male skupiny přilákat mužské přistěhovalců a mají vyšší novorozenců sazby, než skupiny s jedním silný dospělý muž v nejlepších letech (Teichroeb, Wikberg, Badescu, & Sicotte, nepublikované údaje; Obrázek 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

Pokud ekologických omezení model popisující tlaky, které obecně ovlivňují primát velikosti skupiny a případné odchylky od tohoto očekává, že velikost skupiny výsledek od sociálních tlaků, bude to vzrušující doba v této oblasti výzkumu, protože to vyvolává řadu zajímavých otázek. Příklad: Jaká je povaha sociálních tlaků na velikost skupiny? Proč jsou tato sociální omezení přítomna u těchto druhů a nejsou úzce příbuzná? A jaké evoluční síly volí pro tyto sociální tlaky? Velikost skupiny je rozhodující prvek ovlivňující typ sociální organizace druhu přijmout, tedy zkoumání těchto problémů povede k vzrušující vylepšení socioecological modely.
Leave a Reply