8.5: Receptores de tirosina Quinasas (RTKs)
Los receptores de tirosina quinasas median respuestas a un gran número de señales, incluidas hormonas peptídicas como la insulina y factores de crecimiento como el factor de crecimiento epidérmico. Al igual que los GPCR, los receptores de tirosina cinasas se unen a una señal y luego transmiten el mensaje a través de una serie de moléculas intracelulares, la última de las cuales actúa sobre las proteínas diana para cambiar el estado de la célula.
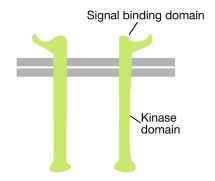
Como su nombre indica, un receptor tirosina cinasa es un receptor de superficie celular que también tiene actividad tirosina cinasa. El dominio de unión de señales del receptor tirosina cinasa se encuentra en la superficie celular, mientras que la actividad enzimática de la tirosina cinasa reside en la parte citoplasmática de la proteína (ver la figura anterior). Una hélice alfa transmembrana conecta estas dos regiones del receptor.
¿Qué sucede cuando las moléculas de señal se unen a los receptores de tirosina quinasas?
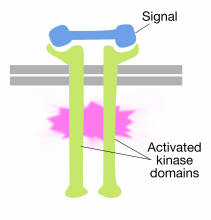
Unión de moléculas de señales a los dominios extracelulares del receptor las moléculas de tirosina cinasa hacen que dos moléculas receptoras se dimericen (se unan y se asocien). Esto acerca las colas citoplasmáticas de los receptores entre sí y hace que la actividad de la tirosina cinasa de estas colas se active. Las colas activadas se fosforilan entre sí en varios residuos de tirosina. Esto se llama autofosforilación.
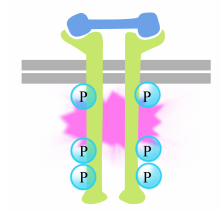
La fosforilación de tirosinas en las colas del receptor provoca el montaje de un complejo de señalización intracelular en las colas. Las tirosinas recién fosforiladas sirven como sitios de unión para las proteínas de señalización que luego transmiten el mensaje a otras proteínas. Una proteína importante que es posteriormente activada por los complejos de señalización en el receptor de tirosina quinasas se llama Ras.
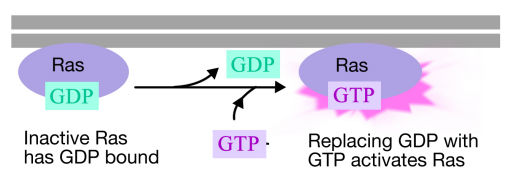
La proteína Ras es una proteína de unión a nucleótidos de guanina monomérica que se asocia con la cara citosólica de la membrana plasmática (de hecho, se parece mucho a la subunidad alfa de las proteínas G triméricas). Al igual que la subunidad alfa de una proteína G, Ras es activa cuando el GTP está unido a ella e inactiva cuando el GDP está unido a ella.Además, al igual que la subunidad alfa, Ras puede hidrolizar el GTP al PIB.
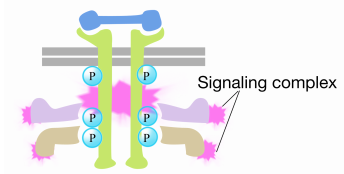
Cuando una señal llega al receptor tirosina quinasa, los monómeros del receptor se unen y fosforilan las tirosinas de los demás, desencadenando el ensamblaje de un complejo de proteínas en la cola citoplasmática del receptor. Una de las proteínas de este complejo interactúa con Ras y estimula el intercambio del PIB unido al Ras inactivo por un GTP. Esto activa el Ras.
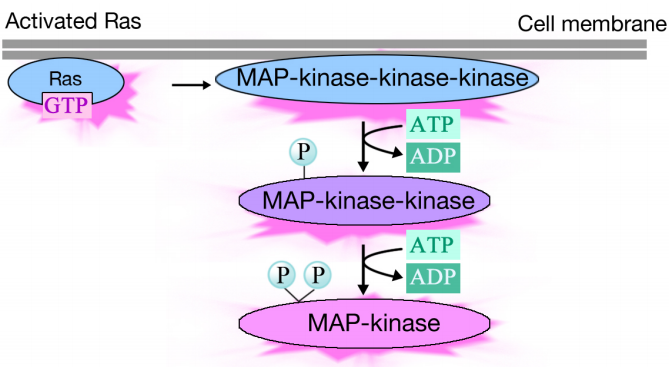
El Ras activado desencadena una cascada de fosforilación de tres proteínas quinasas, que retransmiten y distribuyen la señal. Estas proteínas quinasas son miembros de un grupo llamado quinasas MAP (Proteínas quinasas activadas por Mitógenos). La quinasa final en esta cascada fosforila varias proteínas diana, incluyendo enzimas y activadores transcripcionales que regulan la expresión génica.
La fosforilación de varias enzimas puede alterar sus actividades y desencadenar nuevas reacciones químicas en la célula, mientras que la fosforilación de activadores transcripcionales puede cambiar qué genes se expresan. El efecto combinado de los cambios en la expresión génica y la actividad proteica altera el estado fisiológico de la célula.
Una vez más, siguiendo el camino de la transducción de señales mediada por RTK, es posible discernir el mismo patrón básico de eventos: una señal está unida a los dominios extracelulares de las tirosinas cinasas receptoras, lo que resulta en la dimerización del receptor y la autofosforilación de las colas citosólicas, transmitiendo así el mensaje al interior de la célula.
El mensaje se transmite a través de un complejo de señalización a Ras que luego estimula una serie de quinasas. La quinasa terminal en la cascada actúa sobre las proteínas diana y produce cambios en las actividades de las proteínas y la expresión génica.
Las descripciones anteriores proporcionan un bosquejo muy simple de algunas de las principales clases de receptores y tratan principalmente de los detalles mecanicistas de los pasos por los que las señales recibidas por varios tipos de receptores provocan cambios en las células. Una lección importante para llevar a casa es la similitud esencial de los diferentes caminos.
Otro punto a tener en cuenta es que, si bien hemos observado cada vía individual de forma aislada, una célula, en un momento dado, recibe múltiples señales que desencadenan una variedad de respuestas diferentes a la vez. Las vías descritas anteriormente muestran un grado considerable de» conversación cruzada » y la respuesta a cualquier señal dada se ve afectada por las otras señales que la célula recibe simultáneamente. La multitud de diferentes receptores, señales y sus combinaciones son los medios por los cuales las células son capaces de responder a una enorme variedad de circunstancias diferentes.
Colaboradores
-
Dr. Kevin Ahern y Dra. Indira Rajagopal (Oregon State University)
Leave a Reply