¿Qué Influye en el Tamaño de los Grupos en los que los Primates eligen Vivir?

Una de las principales contribuciones teóricas hechas por el estudio de los primates es la comprensión de lo que influye en la naturaleza de la variación compleja en la estructura y las organizaciones sociales. Detrás de esto hay un conjunto de desarrollos teóricos que examinan por qué los animales viven en grupos en primer lugar y por qué los grupos son de un tamaño particular. Aquí, nos centramos en la pregunta de » ¿qué influye en el tamaño de los grupos en los que los primates eligen vivir?»
Los primates son taxones particularmente útiles para abordar esta pregunta porque el tamaño de sus grupos es muy variable, no solo dentro de las especies, sino también entre ellas. El tamaño intraespecífico del colobo rojo (Procolobus rufomitratus), por ejemplo, varía entre 12 y 150 miembros (Chapman & Chapman 2000a). La variación interespecífica varía de un solo individuo, como se ve en los orangutanes (Pongo sp.) (van Schaik, 1999), a más de 800 miembros de mandrills (Mandrillus sphinx) (Abernethy et al. 2002). Además, dentro de algunas especies, los grupos sociales repetidamente se dividen y se reúnen en subgrupos de diferentes tamaños y combinaciones a lo largo del tiempo (por ejemplo, la organización social de fisión-fusión de monos araña (Ateles sp.) y chimpancés (Pan troglodytes), o la organización multinivel de gelada (Theropithecus gelada) y babuinos Hamadryas (Papio hamadryas, Aureli et al. 2008). Esta variación proporciona la base para que los investigadores desarrollen modelos para investigar los factores ecológicos y sociales del tamaño del grupo.
La agrupación es beneficiosa de varias maneras. Se cree que los individuos en grupos más grandes tienen un menor riesgo de depredación (Hamilton 1971), pueden ser más capaces de encontrar y defender los recursos alimentarios (Cody 1971, Wrangham 1980), y pueden estar protegidos contra amenazas conespecíficas, como el infanticidio de machos extragrupales (Wrangham 1979). Varios investigadores han sugerido que la agrupación confiere beneficios predecibles (Alexander 1974, van Schaik 1983) que las diferencias en el tamaño del grupo pueden explicarse por las desventajas (Wrangham et al. 1993). Se cree que el costo potencial de agrupamiento más ampliamente aceptado es una reducción de la eficiencia de forrajeo. Estar con otros individuos con los mismos requisitos dietéticos significa que los animales se pelean por la comida (competencia de concurso), o un animal en un grupo gana a otro por la comida, por lo que cuando el segundo animal llega a un área simplemente no queda comida (competencia de revueltas, Janson & van Schaik 1988). En ambas situaciones, se cree que la competencia por la comida lleva a los animales a viajar más lejos. La lógica detrás de este argumento es relativamente simple. Los animales deben forrajear en un área que pueda satisfacer sus necesidades energéticas y nutricionales. De ello se desprende que un aumento del tamaño del grupo aumentará el área que debe cubrirse para encontrar suministros de alimentos adecuados. Por lo tanto, los individuos deben viajar más lejos y gastar más energía si están en un grupo grande, que si se alimentan en un grupo más pequeño. Con un aumento en el tiempo de viaje, se acerca un punto en el que la energía gastada en el viaje es demasiado costosa y los grupos más pequeños se vuelven ventajosos. De esta manera, los factores ecológicos pueden influir en los patrones de movimiento y en la eficiencia de la alimentación, limitando así el tamaño de los grupos que pueden explotar eficientemente los recursos alimentarios disponibles. Estas ideas han sido formalizados en lo que se conoce como las limitantes Ecológicas (Modelo de Chapman & Chapman 2000b, Ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
El componente esencial del modelo de restricciones ecológicas es que un aumento en el tamaño del grupo debe conducir a un aumento en la competencia de alimentación dentro del grupo. Es concebible que esto funcione de una manera ligeramente diferente dependiendo de la naturaleza de los recursos utilizados por especies particulares. Con los primates frugívoros, y posiblemente muchos primates folívoros, que se alimentan en parches discretos, típicamente árboles que contienen alimentos o grupos de árboles, los miembros adicionales del grupo pueden agotar los parches más rápido y llevar a un aumento de los rangos de días(Chapman 1988, Snaith & Chapman 2005). Para las especies más insectívoras, cuyos recursos pueden no encontrarse en parches discretos, el viaje continuo a través del dosel en busca de insectos con miembros adicionales del grupo puede conducir a un aumento en la superposición de campos de búsqueda individuales, reduciendo las tasas de encuentro per cápita con los alimentos y, por lo tanto, aumentando el área que se debe buscar (van Schaik et al. 1983).
Por lo tanto, el modelo de restricciones ecológicas sugiere que los factores que afectan la distancia que viajan los animales también deben afectar el tamaño del grupo. Para aquellos animales que normalmente se alimentan de frutas u hojas que pueden agotarse, el tamaño del parche determinaría cuánto tiempo un grupo de un tamaño determinado podría permanecer y alimentarse. Un grupo grande pasaría menos tiempo en un parche de un tamaño dado que un grupo más pequeño, porque agota el parche más rápido: un grupo grande simplemente tiene más bocas que alimentar. Si los animales viajan entre parches una vez que los han agotado, la densidad y distribución de los parches determinarán los costos de viaje incurridos. Cuando los parches de recursos están en una alta densidad o en una distribución agrupada, la distancia al siguiente parche es pequeña, los costos de viaje son bajos y, por lo tanto, los animales pueden formar grupos grandes. En esos momentos, cualquier costo adicional asociado con ser miembro de un grupo grande, como la necesidad de visitar muchos parches, se puede recuperar fácilmente. Por el contrario, cuando los parches de recursos se producen a bajas densidades, la distancia al siguiente parche suele ser grande, los costos de viaje son altos y los animales no pueden permitirse agotar rápidamente los parches y, por lo tanto, los animales forman grupos pequeños.
La organización social de fisión-fusión de monos araña, chimpancés y algunos otros primates (Figura 1) ofrece pruebas útiles del modelo de restricciones ecológicas porque se pueden intentar predecir cambios temporales en el tamaño de los subgrupos a partir de medidas directas del tamaño, la densidad y la distribución de los recursos alimentarios (Chapman 1990, Chapman et al. 1995). Para los monos araña y los chimpancés, se desarrolló un modelo de regresión múltiple para mostrar que el tamaño, la densidad y la distribución de los parches de alimentos medidos cada mes eran un buen predictor del tamaño de los subgrupos. Es difícil representar visualmente una regresión múltiple, pero los métodos que se utilizaron para los monos araña se muestran en la Figura 2. Los monos araña tienen un tipo de organización social de fisión-fusión muy flexible, y en el Parque Nacional Santa Rosa, Costa Rica, el tamaño de los subgrupos puede variar de 1 a 35 individuos, pero en promedio se encuentran alrededor de 5 individuos viajando juntos. Para esta población, el 50% de la varianza en el tamaño medio mensual del subgrupo se puede predecir a partir de medidas relativamente crudas del tamaño, la densidad y la distribución de los parches de alimentos (Chapman 1990).

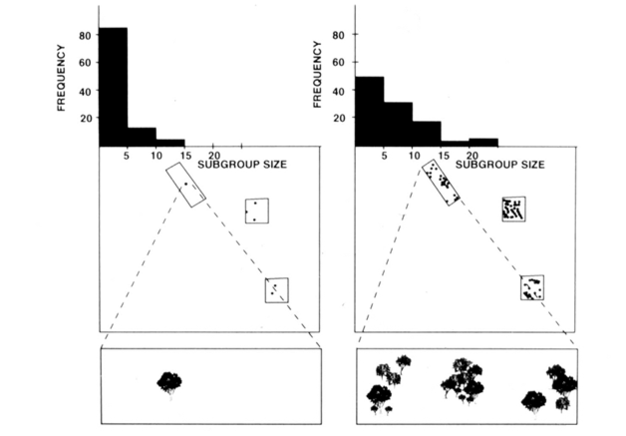
Además de las especies mencionadas anteriormente, la aplicabilidad del modelo de restricciones ecológicas ha sido generalmente apoyada con investigaciones provenientes de una variedad de especies (Snaith & Chapman 2005) y situaciones (por ejemplo, modelos generales, Wrangham et al. 1993, y asociaciones de especies mixtas, Chapman & Chapman 2000c). Consideramos que, dado el amplio apoyo que ha recibido este modelo, ha llegado el momento de que sea razonable sugerir que las especies deben ajustarse típicamente a las expectativas del modelo de limitaciones ecológicas. Esto no significa que todas las especies se conformen, y por lo tanto se convierte en un momento emocionante para la investigación en este campo porque los investigadores pueden buscar excepciones al modelo. Sospechamos que las especies que no se ajustan lo harán porque han adoptado estrategias sociales que van en contra de las expectativas ecológicas. Esto proporciona un «patrón» para evaluar la importancia potencial de varias estrategias sociales (es decir, cuanto más se desvía de las expectativas del modelo ecológico, más importantes son las presiones selectivas sociales).
Proporcionemos brevemente dos ejemplos de nuestra investigación anterior. Las hembras de chimpancé eran más solitarias que las hembras de mono araña, sin embargo, ambas tienen organizaciones sociales de fisión-fusión similares. Además, si bien pudimos predecir con precisión el número de chimpancés machos y subadultos en un subgrupo en función de las condiciones ecológicas, no pudimos predecir el número de chimpancés femeninos en estos subgrupos (Chapman et al. 1995). Incluso cuando los recursos eran extremadamente abundantes y casi todos los machos estaban en un subgrupo grande, las hembras rara vez ingresaban a grupos, lo que sugiere que el costo de estar en un subgrupo más grande superaba cualquier beneficio, como evitar depredadores. Esto es inesperado desde la perspectiva de que los bebés de estas hembras serían la clase de edad/sexo más amenazada por la depredación. Una hipótesis comprobable para explicar estas observaciones es que la naturaleza de las coaliciones en estas dos especies puede influir en los beneficios de la pertenencia a un grupo. La evidencia sugiere que, a diferencia de los chimpancés, las hembras de mono araña forman coaliciones que a menudo operan para permitir a los miembros de la coalición acceso exclusivo a los alimentos. Por lo tanto, los monos araña, cuando se unen a individuos particulares, pueden aumentar su acceso a la comida al excluir a otros, pero para las hembras de chimpancé, este no es el caso (Chapman et al. 1995).
Identificamos otra desviación interesante, potencialmente causada por factores sociales que afectan los costos y beneficios de estar en un grupo, y esto se refiere a los monos colobo rojo y colobo blanco y negro (aka. guerezas, Colobo guereza) en el Parque Nacional de Kibale, Uganda. Los colobos rojos forman grandes grupos con un promedio de 65 individuos(25-127 individuos, Snaith et al. 2008), mientras que los guerezas viven en pequeños grupos con un promedio de 6,5 individuos (4-11 individuos, Harris & Chapman 2007). A pesar de esta diferencia, hay una gran similitud en las plantas que comen las dos especies. Si las condiciones ecológicas fueran responsables de la diferencia en el tamaño del grupo entre las dos especies de colobines, se esperaría que sus dietas fueran diferentes y la densidad de los árboles alimentarios fuera menor en las áreas de distribución de las guerezas, ya que tienen el tamaño del grupo más pequeño. Sin embargo, sus dietas son muy similares; la superposición dietética entre un colobo rojo y un grupo de guereza que tenía su rango de hogar completamente dentro del rango de hogar del grupo de colobo rojo fue de 43.2%, mientras que para los dos grupos vecinos de colobo rojo, la dieta se solapó solo en un 37,3% (Chapman et al. 2002). Parece que el éxito reproductivo de la guereza femenina se maximiza en grupos pequeños y medianos, ya sea porque los grupos más grandes experimentan tasas más altas de tomas e infanticidio o más competencia de alimentación (ver evidencia de competencia de revueltas en el folívoro Trachypithecus phayrei y Semnopithecus sp. , pero falta de evidencia de que guerezas haya agotado los parches de comida). Hemos sido testigos de una serie de secuestros de hombres y asesinatos de niños en las guerezas (Onderdonk 2000)-véase Teichroeb & Sicotte (2008) para una evaluación de la teoría sobre el infanticidio en colobines. Lo que no está claro es cómo el riesgo de infanticidio limitaría el tamaño de los grupos de guereza, cuando aparentemente tienen acceso a una alta densidad de alimentos que debería permitir la formación de grupos de hombres múltiples. En varias especies, los grupos de hembras más grandes son más un objetivo para los machos inmigrantes, y esta situación puede aumentar las tasas de infanticidio, por ejemplo, Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). Sin embargo, se espera que el infanticidio sea menos frecuente si un grupo contiene múltiples machos activos reproductivamente, porque la inmigración masculina y la posterior toma de posesión violenta y el infanticidio son menos probables cuando el recién llegado se enfrenta a varios posibles hijos de futuros bebés (Janson & van Schaik 2000). Las guerezas parecen tener amplios recursos alimentarios para formar grupos más grandes (Tombak et al. 2012), lo que disminuiría el riesgo de infanticidio, pero no forman grupos grandes. Es posible que la naturaleza de las interacciones hombre-hombre en guerezas haga que los grupos grandes sean inestables. Este parece ser el caso del colobo ursino (Colobus vellerosus), donde los grupos de hombres múltiples atraen inmigrantes masculinos y tienen tasas de infanticidio más altas que los grupos con un hombre adulto fuerte en su mejor momento (Teichroeb, Wikberg, Badescu, & Sicotte, datos no publicados; Figura 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

Si el modelo de restricciones ecológicas describe las presiones que generalmente influyen en el tamaño del grupo de primates y si las desviaciones de este tamaño esperado del grupo resultan de presiones sociales, será un momento emocionante en esta área de investigación porque plantea una serie de preguntas interesantes. Por ejemplo: ¿Cuál es la naturaleza de las presiones sociales sobre el tamaño del grupo? ¿Por qué estas limitaciones sociales están presentes en estas especies y no están estrechamente relacionadas? ¿Y cuáles son las fuerzas evolutivas que seleccionan para estas presiones sociales? El tamaño del grupo es un elemento crítico que influye en el tipo de organización social que adoptan las especies, por lo que el examen de estos temas conducirá a mejoras emocionantes de los modelos socioecológicos.
Leave a Reply