桑実胚の開発
Google翻訳-言語を選択リストから以下の通りです(これは新たな外部ページ)
العربية|català|中文|中國傳統的|ィチュ-フランセ|ドイツ|עִברִית|हिंदी|インドネシア語|italiano|english|한국어|မြန်မာ|フィリピン|Polskie|português|ਪੰਜਾਬੀਦੇ|Română| русский|Español|スワヒリ語|Svensk|ไทย|Türkçe|اردو|ייִדיש/TiếNg ViệTこれらの外部の翻訳は自動化されていない場合も正確なものでなければならない。 (もっと? 翻訳について)
はじめに
(ラテン語、桑実胚=桑)細胞が急速に有糸分裂して”桑”の外観を持つ細胞(12-15細胞)の固体塊を生成する受精後の開発の初期段階。 この段階に続いて、この細胞塊胚盤胞の段階で空洞が形成される。
桑実胚形成前の重要なイベントは、8細胞胚が細胞形態および細胞-細胞接着の変化を受け、この細胞の固体ボールの形成を開始する”圧縮”である。
ヒトでは、受精後の第一週(GA第3週)の最初の日に発生し、カーネギー第2期と記載されている。 この段階に続いて、胚盤胞の形成を定義する空洞、胚盤胞の形成が続く。p>

アート着床前胚盤胞生検
|
|
アート着床前胚盤胞生検生殖補助技術では、桑実胚の段階は、単一の細胞(胚盤胞)を除去し、そのdna上の遺伝子診断を行うことによって、最も初期の出生前診断検査の一つを行うこ
|
分子-マウスでは、桑実胚期から胚盤胞期への移行中に、内細胞塊(ICM)および栄養外胚葉(TE)の分化は、Hippo経路によって調節されることが示されている。Morulaリンク:morula|カーネギーステージ2/有糸分裂|胚盤胞|受精|週1/講義-週1|カテゴリ:カーネギーステージ2|カテゴリ:Morula
いくつかの最近の知見
- カバ経路メンバー YAP1とLATS2転写産物をダウンレギュレートする効果ブタ胚の分化に関与する初期の開発と遺伝子….. 本研究では、我々はカバ経路メンバー yes関連タンパク質1(YAP1)と大腫瘍サプレッサー2(LATS2)とRNA干渉を用いてブタ着床前開発中にこれらの遺伝子の機能の遺伝子発現パターンを調べた。 YAP1とLATS2mRNAレベルの両方がin vitroで成熟した卵母細胞と1細胞段階の胚で高く示され、開発と徐々に落ちた。 YAP1核局在は、桑実胚および胚盤胞の段階で検出された。 YAP1またはLATS2のいずれかのダウンレギュレーションは、ブタの着床前の開発を阻害し、POUクラス5homeobox1(OCT-4)とSRY関連HMGボックス遺伝子2(SOX2)、icm/TE分化に必 まとめると、YAP1とLATS2は、ブタの着床前の開発のために不可欠であり、それはカバ経路がブタICM/TE分離に重要な役割を持っている可能性があります。”
- 開発および成体組織におけるヒトおよびマウスhomeobox遺伝子発現の多様性”morulaへの8細胞、生殖組織または胚の外で検出できない12個のユテリアン特異的homeobox遺伝子(RHOXF2、RHOXF2B、CPHX1、CPHX2、DPRX、LEUTX、TPRX1、TPRX2、ARGFX、NANOGNB、DUXA、duxb)。 Hox

この表は、リストされた”検索用語”テキストリンクを使用して、外部PubMedデータベースの自動コンピュータ検索を可能にします。
- この検索では、元のPubMed拡張機能が無効になっているため、手動リンクが必要になりました。
- 表示された参考文献のリストは、コンテンツまたは関連性に基づく資料の編集上の選択を反映していません。
- 参照は、実際のページ表示日に基づいてこのリストにも表示されます。
コンテンツページの残りの部分と関連するディスカッションページ(出版年のサブ見出しの下にリストされている)に記載されている参照には、関連性と可用性の両方に基づいていくつかの編集選択が含まれています。
もっと?
もっと? 参考文献/ディスカッションページ|ジャーナル検索|2019参考文献/2020参考文献
検索語:Morula Development|Morula Compaction|Morula Blastomere Division|Morula Hippo/
これらの論文はもともといくつかの最近の調査結果表に登場しましたが、そのリストが長くなるにつれて、この折りたたみ可能な表にシャッフルされました。
この現在のページの年および参考文献によってリストされている他の参考文献については、議論ページも参照してくださ
- Par-aPKC依存性および独立したメカニズムは、16細胞期マウス胚における細胞極性、カバシグナル伝達、および細胞位置決めを協調的に制御する”着床前 32以上の細胞を持つ初期の胚盤胞では、Par-aPKCシステムは、apicobasal軸に沿って外側の細胞の分極を制御し、細胞の極性はHippoシグナル伝達を抑制する。”Hippo/mouse
- 胚ゲノム活性化前のヒト胚の非侵襲的イメージングは、胚盤胞の段階への開発を予測する”我々は、タイムラプス画像解析と遺伝子発現プロファイリングを相関する着床前のヒト胚の開発の研究を報告しています。 体外受精(IVF)から接合体の大規模なセットを調べることにより、我々は胚盤胞期への進行の成功は、胚ゲノム活性化(EGA)の前に、受精後2日目までに三つの動”
- blastomere single-cell cDNA analysisによる5-to8-cell stage human胚の機能的ゲノミクス”5-to8-cell human胚から四十九blastomeresは、高密度マイクロアレイ分析のためのテンプレートを提供するために、効率的な単一細胞cDNA増幅プロトコルに従って調査されている。 以前に記載されたマーカー、内部細胞塊(ICM)(n=120)、幹(n=190)と栄養外胚葉(TE)(n=45)の特徴を分析し、46遺伝子のハウスキーピングパターンが確立されました。 …要約すると、5-8-細胞段階のヒト胚のグローバル単細胞cDNA増幅マイクロアレイ分析は、胚盤胞の運命はICMまたはTEにコミットされていないことを明ら”
映画
|
|
Zygote Mitosis Page | Play
|
Early Division Page | Play
|
Parental Genomes Page | Play
|
Mouse Blastocyst Page | Play
|
Movies
Compaction
- E-cadherin mediated adhesion initiates at compaction at the 8-cell stage
- regulated post-translationally via protein kinase C and other signalling molecules
Blastomere Division

ヒト胚盤胞の発達に関するin vitro研究では、最初に急速に分裂する胚盤胞が胚盤胞の段階に発達する可能性が高いことが示された。
マウスでの最近の研究では、8から16の細胞段階移行における細胞分裂中に有糸分裂紡錘体の特異的な配向がなかったことが示された。 これは、8細胞段階では所定の切断パターン(プレパターン)がなく、有糸分裂中の細胞の切り上げの程度によってのみ調節されることを示唆している。 このようなワームCなどの他の種では、。elegansとascidiansは、接合体の段階から紡錘体の向きの特定のパターンを持っています。
モデルヒト桑実胚発生
次の図は、受精後の第1週のin vitroヒト発達のビデオおよび遺伝子解析を用いた最近の研究からのものです。p>
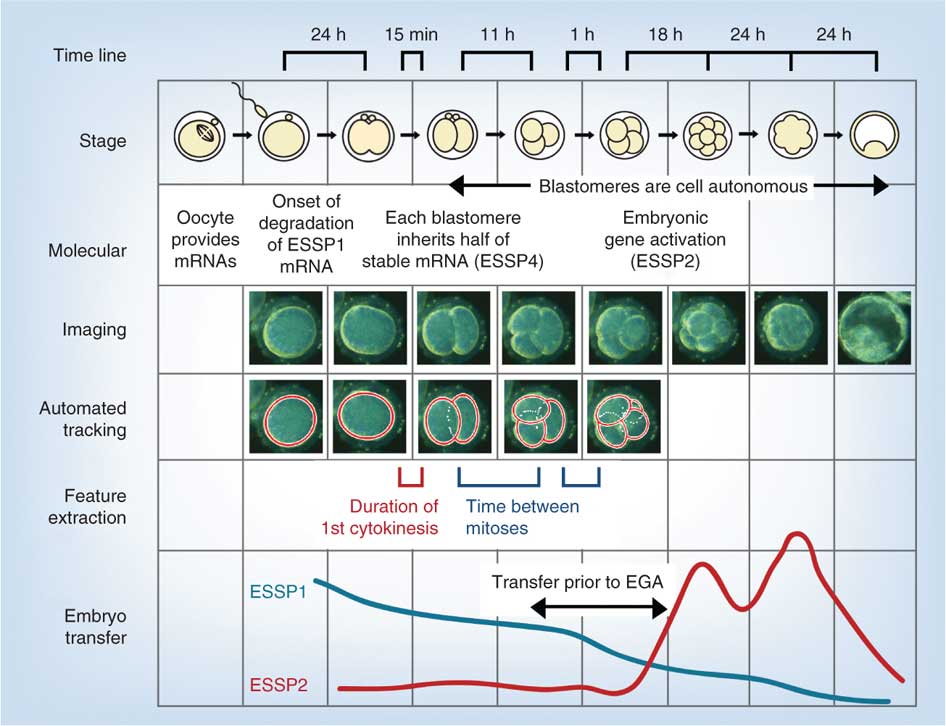
- EGA-胚ゲノム活性化
- ESSP-胚期特異的パターン、四つのユニークな胚期特異的パターン(1-4)
リンク: 伝説の図
他の種のMorulas
マウスMorula
マウスでは、morulaから胚盤胞への移行期に、内細胞塊(ICM)と栄養外胚葉(TE)の分化がHippo経路によって調節されることが示されている。
-

4 cell morula stage development
-

Sox2 expression
-

Early gene expression
-

Early gene expression
-

Early gene expression
-

Early リンク:マウス開発
ウニモルーラ

ウニ初期胚切断パターン(SDBギャラリー画像)
リンク:ウニ開発
ウシMorula
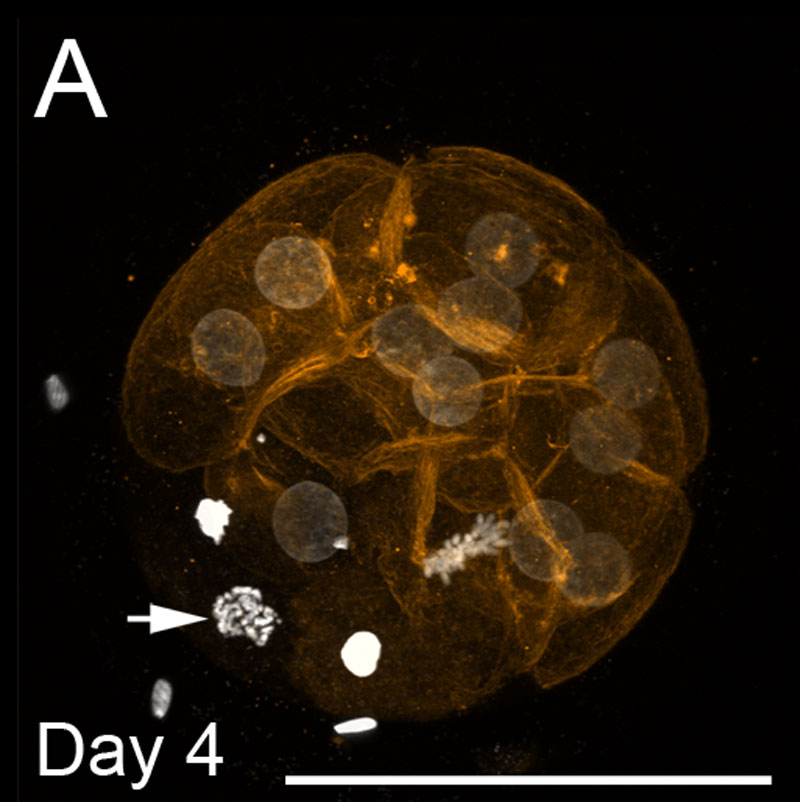
ウシMorula
- 画像は、4日目のDNA染色(白)とf-アクチンフィラメント(オレンジ)を示しています。 スケールバーは100μ mを表します。
- 淡い染色丸い核は間期にある。
- 矢印は前期で単一の核を示しています。
- 中期には単一の核が見られる。
- 凝縮した明るい核はアポトーシスである。
リンク:ウシの発達/有糸分裂
モルーラ生検

コンパクトmorula段階胚の生検
- (A)生検前のコンパクトmorula段階胚。
- (B–G)生検のステップ。
- (H)生検後2時間の胚。リンク:出生前診断|生殖補助技術
- 1.0 1。1Zhang P,Zucchelli M,Bruce S,Hambiliki F,Stavreus-Evers A,Levkov L,Skottman H,Kerkelä E,Kere J&Hovatta O.(2009). ヒト移植前発達のトランスクリプトームプロファイリング。 PLoS ONE,4,e7844. PMID:19924284DOI.
- Milachich T.(2014)。 赤ちゃんの健康状態の潜在的な予測因子としての着床前および出生前の遺伝子スクリーニングおよび非侵襲的検査の新しい進歩。 Biomed Res Int,2014,306505. 質問番号:24783200
- 3.0 3.1平手Y,平原S,井上K,清成H,丹羽H&佐々木H.(2015). Par-aPKC依存性と独立したメカニズムが協力して細胞極性、カバのシグナル伝達、および16細胞段階のマウス胚における細胞の位置決めを制御します。 デヴ 成長は異なる。 , 57, 544-56. 質問番号:26450797
- 4.0 4.1 4.2江村N,斉藤Y,三浦R&沢井K.(2020). ヒッポ経路メンバー YAP1とLATS2転写産物の早期開発とブタ胚の分化に関与する遺伝子発現をダウンレギュレートの効果。 セル再プログラム、、。 質問番号:32150685
- DUNWELL TL&オランダPW。 (2016). 発達および成体組織におけるヒトおよびマウスhomeobox遺伝子発現の多様性。 BMC Dev. バイオル , 16, 40. 質問番号:27809766
- 6.0 6.1Wong CC,Loewke KE,Bossert NL,Behr B,De Jonge CJ,Baer TM&Reijo Pera RA. (2010). 胚ゲノム活性化前のヒト胚の非侵襲的イメージングは、胚盤胞の段階への発達を予測する。 ナット バイオテクノール , 28, 1115-21. PMID:20890283DOI.
- Galán A,Montaner D,Póo ME,Valbuena D,Ruiz V,Aguilar C,Dopazo J&Simón C.(2010). Blastomere単細胞cDNA分析による5-8細胞段階のヒト胚の機能ゲノミクス。 PLoS ONE,5,e13615. PMID:21049019DOI.
- 8.0 8.1Dard N,Louvet-Vallée S&Maro B.(2009). マウス胚における8-16細胞段階移行中の有糸分裂スピンドルの向き。 PLoS ONE、4、e8171。 PMID:19997595DOI.
- Fenwick J,Platteau P,Murdoch AP&Herbert M.(2002). 授精から最初の切断までの時間は、in vitroでのヒト着床前胚の発達能力を予測する。 ハム リプロド , 17, 407-12. PMID:11821286
- Leidenfrost S,Boelhauve M,Reichenbach M,Güngör T,Reichenbach HD,Sinowatz F,Wolf E&Habermann FA. (2011). 哺乳類の着床前発達における細胞停止および細胞死:ウシモデルからの教訓。 PLoS ONE,6,e22121. 質問番号:21811561
- Zakharova EE,Zaletova VV&Krivokharchenko AS. (2014). ヒト桑実胚の生検:PGSによる215IVF/ICSIサイクルの結果。 PLoS ONE,9,e106433. PMID:25191937DOI.
レビュー
Coticchio G,Lagalla C,Sturmey R,Pennetta F&Borini A.(2019). 謎のmorula:開発のメカニズム、細胞の運命の決定、自己補正と芸術のための含意。 ハム リプロド 更新、25、422-438。 PMID:30855681DOI.
記事
Bessonnard S,Mesnard D&Constam DB. (2015). PC7と関連するプロテアーゼFurinとPace4は、胚盤胞形成中にE-カドヘリン機能を調節します。 J.Cell Biol. , 210, 1185-97. PMID:26416966DOI.
Dzamba BJ,Jakab KR,Marsden M,Schwartz MA&DeSimone DW. (2009). カドヘリン接着、組織張力、および非カノニカルWntシグナル伝達は、フィブロネクチンマトリックス組織を調節する。 デヴ セル、16、421-32。 PMID:19289087DOI.
Santos J,Pereira CF,Di-Gregorio A,Spruce T,Alder O,Rodriguez T,Azuara V,Merkenschlager m&Fisher AG. (2010). 多能性および胚外幹細胞のエピジェネティックおよびリプログラミング特性の違いは、開発中のマウス胚における重要な初期のイベントとしてクロマチンのリモデリングを関与させます。 エピジェネティクス-クロマチン,3,1. PMID:20157423DOI.用語集リンク集:A/B|C|D/E/F/G|H|I|J|K|L|M|N|O|P|Q|R|S|T|U|V|W|X|Y|Z|Numbers|Symbols|Term Link
用語集:a|B|C|D|E|F/G/H/I/J/K/L/M/N/O/P/Q/R/S/T/U/V/W/X/Y/Z/Numbers/Symbols/Term Link
用語集:a/B/C/D/E/F/G/H/I/J/K/L/M/N/O/P/Q/R/S/T/U/V/W/X/Y/Z/Numbers/Symbols/Term Link
用語集:このページを引用:Hill、m.a.(2021、3月24日)発生学morulaの開発。 から取得https://embryology.med.unsw.edu.au/embryology/index.php/Morula_Development
Leave a Reply