細菌の染色体:それらはすべて単一で円形ですか?
長年にわたり、すべての細菌は、線形染色体を有する真核生物と比較して、単一の円形染色体を有すると考えられていた。 おそらく、これは、当時研究されていた細菌株の限られたサンプルと、細菌染色体を調べるために利用可能な配列決定技術によるものでした。 しかし、この信念は、1989年に複数の染色体および/または線状の染色体を有する細菌が発見されたときに反証された。 P>
単一および環状染色体
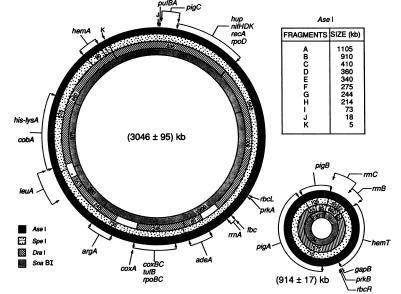
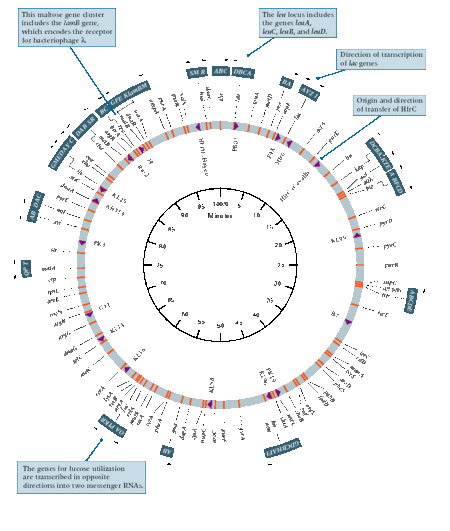 大腸菌の環状染色体マップ
大腸菌の環状染色体マップ1963年に、いくつかの研究者によって行われた研究では、大腸菌のゲノムは単一の環状染色体に組織化されてい 染色体DNAの構造の証拠は、オートラジオグラフィー、電子顕微鏡および蛍光顕微鏡を用いたDNAの動画像によって達成された画像によって示された。 ケアンズは、大腸菌の染色体全体の画像を取得した最初の研究者でした。 使用された技術は、大腸菌の染色体を水素の放射性同位体であるトリチウム標識チミジンを用いて標識するオートラジオグラフィーであった。 しかし、染色体の大きさは可変であり、検出された円形の形態の頻度は低かった。 Hfr抱合に基づくさらなる実験は、大腸菌が環状染色体を有することを説得力のある実証している。
大腸菌の環状染色体の公開されたデータは、多くの研究者によって迅速に採用されました。 したがって、大腸菌は染色体複製を調査するための主要なモデルとして広く使用されるようになった。 P>
複数の染色体
線形染色体
1970年に、線形染色体の最初の証拠が発見されましたが、科学的技術の利用可能性が限られており、細菌の染色体の循環性の強い信念は、多くの人々がずっと後までこの考えを受け入れなかったほど説得力がありました。 1989年までに、パルスフィールドゲル電気泳動法が開発され、spirochete Borrelia burgdorferiがr.sphaeroidesのような分離された断片を接続することによって絞り込みの同様のプロセスを使用して線形染色体を持っていることを確認するために制限消化と組み合わせて使用された。 B. burgdorferiは、線状染色体が原核生物に存在することを示した最初の細菌であり、染色体のサイズは約1.0Mbであることが判明した。 この染色体組織は真核生物の染色体組織に匹敵する。
線状染色体の問題
原核生物における環状染色体と比較して、線状染色体の存在と使用には二つの問題が生じます。 第一に、遊離二本鎖DNA末端は、細胞内ヌクレアーゼがそれらを分解しないように、何らかの保護を持たなければならない。 第二に、直鎖状DNA分子の末端であるテロメアは、DNA複製のために異なるプロセスを必要とする。 保護のために、観察されたテロメアの2つのタイプがあります。 最初のものは回文ヘアピンループと呼ばれ、利用可能な自由な二本鎖端がない。 他のタイプは染色体が5’端に結合する蛋白質を含んでいるinvertronテロメアです。 これはStreptomycesの線形染色体で見つけることができます。 直鎖状DNA分子の末端のDNA複製に関しては、そのメカニズムについてはあまり知られていない。
線状染色体の他の所見
s.griseusは、抗生物質、ストレプトマイシンを産生することが知られているグラム陽性細菌です。 この細菌にまた制限の消化と共に脈拍分野のゲルの電気泳動によって識別される線形染色体があります。 線状染色体が同定された他のStreptomyces種もある。
Agrobacterium tumefaciens
パルスフィールドゲル電気泳動を用いて、a.tumefaciensゲノムを分析し、一つの線状染色体と一つの環状染色体を同定した。 線状染色体のサイズは2100Kbであり、これは3000Kbの円形染色体よりも小さい。 アグロバクテリウム種は、二つの円形染色体を持つことが知られているメンバーを持っているブルセラ種と近い16s rRNA配列関係を持っています。
複数の染色体の他の所見
B.melitensisはグラム陰性のcoccobacillus細菌であり、二つの円形染色体を有する。 2つの染色体サイズは2117Kbと1178Kbです。 2つの環状染色体を持つ他のブルセラ種には、B.suis biovar1、B.suis biovar2、B.suis biovar4、B.abortusおよびB.ovisが含まれる。
Paracoccus denitrificans
P.denitrificansは、パルスフィールドゲル電気泳動によって同定された三つの円形染色体を有する特殊な細菌である。 染色体のおおよそのサイズは2、1.1、および0.64Mbです。
ビブリオ種
複数の染色体を持つ他の多くの細菌がありますが、ビブリオ種は2つの円形の染色体を持つメンバーを持っています。
ビブリオ種
これらはV.cholerae、V.parahaemolyticus、V.vulnificusおよびV.fluvialisを含んでいます。
Summary of Currently Known Bacterial Chromosome Organizations
| Bacteria | Chromosome Organization |
|---|---|
| Agrobacterium tumefaciens | One linear and one circular |
| Bacillus subtilis | Single and circular |
| Bacillus subtilis | Single and linear |
| Borrelia burgdorferi | Two circular |
| Brucella abortus | Two circular |
| Brucella melitensis | Two circular |
| Brucella ovis | Two circular |
| Brucella suis biovar 1 | Two circular |
| Brucella suis biovar 2 | Two circular |
| Brucella suis biovar 4 | Two circular |
| Escherichia coli | Single and circular |
| Paracoccus denitrificans | Three circular |
| Pseudomonas aeruginosa | Single and circular |
| Rhodobacter sphaeroides | Two circular |
| Streptomyces griseus | Linear |
| Vibrio cholerae | Two circular |
| Vibrio fluvialis | Two circular |
| Vibrio parahaemolyticus | Two circular |
| Vibrio vulnificus | Two circular |
(1) Allardet-Servent, A., Michaux-Charachon, S., Jumas-Bilak, E., Karayan, L. and Ramuz, M. “Presence of One Linear and One Circular Chromosome in the Agrobacterium tumefaciens C58 Genome.” Journal of Bacteriology, 1993, 175(24): 7869-7874
(2) Baril, C., Richaud, C., Baranton, G. and Saint-Girons, I. “ボレリア-ブルグドルフェリの線形染色体。”とコメントしている。 1 9 8 9,1 4 0:5 0 7−5 1 6(3)Bendich,A.”Biochimie,2001,83:177-186
(4)Cairns,J.”オートラジオグラフィーによって見られる細菌染色体およびその複製様式。”J.Mol. バイオル(5)Del Vecchio,V.G.,Kapatral,V.,Elzer,P.,Patra,G.and Mujer,C.V.”The Genome of Brucella melitensis.”獣医微生物学,2002,90:587-592
(6)Egan,E.S.,Fogel,M.A.and Waldor,M.K.”分割されたゲノム: 複数の染色体を持つ原核生物の細胞周期を交渉する。”分子微生物学, 2005, 56(5): 1129-1138(7)Griswold,A.”Genome Packaging in Prokaryotes:The Circular Chromosome of e.coli.”Nature Education,2008,1(1)
(8)Hinnebusch,J.and Tilly,K.”Linear Plasmids and Chromosos in Bacteria.”分子微生物学, 1993, 10(5): 917-922
(9)Jumas-Bilak,E.,Michaux-Charachon,S.,Bourg,G.,O’Callaghan,D.and Ramuz,M.”Brucella属における染色体数とゲノム再配列の違い。”モル… ミクロビオール, 1998, 27(1): 99-106
(10)Lezhava A.,Mizukami,T.,Kajitani,T.,亀岡,D.,Redenbach,M.,Shinkawa,H.,Nimi,O.and Kinashi,h.”Stremptomyces griseusの線形染色体の物理マップ。”細菌学ジャーナル, 1995, 177(22): 6492-6498
(11)Suwanto,A.and Kaplan,S.”Rhodobacter sphaeroidesにおける染色体移動:二つのユニークな円形染色体のためのHfr形成と遺伝的証拠。”細菌学ジャーナル, 1992, 174(4): 1135-1145
(12)Suwanto,A.and Kaplan,S.”Rhodobacter sphaeroides2.4.1ゲノムの物理的および遺伝的マッピング: 二つのユニークな円形染色体の存在。”細菌学ジャーナル, 1989, 171(11): 5850-5859
(13)Ussery,D.”バクテリオファージ遺伝学.”2000,http://www.cbs.dtu.dk/staff/dave/roanoke/genetics980311.html
(14)Volff,J.N.And Altenbuchner,J.”A New Beginning with New Ends:Linearisation of Circular Chromosomes during Bacterial Evolution.”FEMS Microbiology Letters,2000,186:143-150
(15)Winterstein,C.and Ludwig,B.”呼吸複合体をコードする遺伝子は、Paracoccus denitrificansゲノムの三つの染色体すべてにマップする。”アーチ。 ミクロビオール, 1998, 169(4): 275-281
(16)山一,Y.,飯田,T.,Park,K.S.,Yamamoto,K.and Honda,T.”腸炎ビブリオのゲノムの物理的および遺伝的地図:ビブリオ種における二つの染色体の存在。”モロヘイヤ。, 1999, 31(5): 1513-1521
Leave a Reply