霊長類が生きることを選択するグループのサイズには何が影響しますか?

霊長類は、そのグループのサイズが非常に可変であるため、この質問に対処するために特に有用な分類群であるだけでなく、種間でも。 例えば、赤いコロバス(Procolobus rufomitratus)の種内グループのサイズは、12と150のメンバーの間で変化します(Chapman&Chapman2000a)。 種間変異は、オランウータン(Pongo sp.(van Schaik1 9 9 9)、mandrillus sphinx(Mandrillus sphinx)における8 0 0人以上のメンバーに(Abernethy e t a l. 2002). さらに、いくつかの種の中で、社会集団は、時間の経過とともに異なる大きさおよび組み合わせのサブグループに繰り返し分割および再結合する(例:fission-fusion social organization of spider monkeys(Ateles sp. チンパンジー(Pan troglodyts)、またはgelada(Theropithecus gelada)およびHamadryasヒヒの多レベル組織(Papio hamadryas,Aureli e t a l. 2008). この変化は、研究者がグループサイズの生態学的要因と社会的要因の両方を調査するためのモデルを開発するための基礎を提供する。
グループ化はいくつかの点で有益です。 より大きなグループの個体は、捕食のリスクが低下し(Hamilton1971)、食料資源を見つけて防御することができ(Cody1971、Wrangham1980)、グループ外の男性による幼児化のような同種の脅威から保護される可能性があると考えられている(Wrangham1979)。 様々な研究者は、グループ化がそのような予測可能な利点を与えることを示唆している(Alexander1974、van Schaik1983)、グループサイズの違いは欠点によって説明できる(Wrangham et al. 1993). グループ化の最も広く受け入れられている潜在的なコストは、採餌効率の低下であると考えられている。 同じ食事の要件を持つ他の個人と一緒にいることは、動物が食べ物の上で戦う(コンテストの競争)、またはグループ内のある動物が食べ物に別の動物を打つ これらの状況の両方では、食品上の競争は、動物が遠くに移動する必要があることにつながると考えられています。 この議論の背後にある論理は比較的単純です。 動物は彼らの精力的な、栄養の条件を満たすことができる区域に飼料を与えなければならない。 グループサイズの増加は、適切な食料供給を見つけるためにカバーされなければならない領域を増加させることになる。 したがって、個人は、より小さなグループで餌を食べる場合よりも、彼らが大きなグループにいるならば、さらに移動し、より多くのエネルギーを消費しなければなりません。 旅行に費やされる時間の増加に伴い、旅行に費やされるエネルギーがあまりにも高価であり、より小さなグループが有利になる点が近づいている。 このようにして、生態学的要因が移動パターンと採餌効率に影響を与え、利用可能な食料資源を効率的に利用できるグループの規模を制約する可能性が これらのアイデアは、生態学的制約モデルとして知られるようになったもので形式化されています(Chapman&Chapman2000b、Ganas&Robbins2005、Snaith&Chapman2007、Teichroeb&sicotte2009)。
生態学的制約モデルの本質的な要素は、グループサイズの増加は、グループ内の摂食競争の増加につながる必要があるということです。 これは、特定の種によって使用される資源の性質に応じてわずかに異なる方法で動作することが考えられる。 質素な、そしておそらく多くのfolivorous霊長類では、離散的なパッチ(典型的には食料品を持つ木または木の塊を有する木)を食べるが、追加のグループメンバーは、パッチをより速く枯渇させ、日の範囲を増加させる可能性がある(Chapman1988,Snaith&Chapman2005)。 資源が離散的なパッチとして発生しない可能性のあるより多くの食虫植物種については、追加のグループメンバーを持つ昆虫を探してキャノピー全体を連続的に移動すると、個々の検索フィールドの重複が増加し、一人当たりの食物との遭遇率が減少し、したがって検索されなければならない面積が増加する可能性がある(van Schaik et al. 1983).
だから、生態学的制約モデルは、動物が移動する距離に影響を与える要因もグループのサイズに影響を与えるべきであることを示唆しています。
通常、枯渇する可能性のある果物または葉を食べる動物の場合、パッチのサイズは、所与のサイズのグループがどれくらい長く滞在して餌を与えるこ それはより速くパッチを枯渇させるので、大規模なグループは、小さなグループよりも、特定のサイズのパッチでより少ない時間を費やすだろう—大規模なグ 動物がそれらを枯渇させた後にパッチ間を移動する場合、パッチの密度および分布は、発生した移動コストを決定する。 資源パッチが高密度または凝集分布にある場合、次のパッチまでの距離は小さく、移動コストは低く、動物は大きなグループを形成することができます。 そのような時には、多くのパッチを訪問する必要性など、大規模なグループのメンバーであることに関連する追加コストを簡単に回収することができま 対照的に、資源パッチが低密度で発生すると、次のパッチまでの距離は一般的に大きく、移動コストが高く、動物はパッチを急速に枯渇させる余裕がな
クモザル、チンパンジー、およびいくつかの他の霊長類の核分裂融合社会組織(図1)は、食物資源の大きさ、密度、および分布の直接測定からサブグループサ 1995). クモザルとチンパンジーのために、毎月測定された食品パッチのサイズ、密度、および分布がサブグループサイズの良い予測因子であることを示すために重回帰モデルが開発されました。 重回帰を視覚的に表現することは困難ですが、クモザルに使用された方法を図2に示します。 クモサルは非常に柔軟な核分裂融合型の社会組織を持っており、コスタリカのサンタローザ国立公園では、サブグループのサイズは1人から35人までの範囲であるが、平均して約5人が一緒に旅行していることが分かっている。 この集団では、平均月次サブグループサイズの分散の50%は、食品パッチのサイズ、密度、および分布の比較的粗雑な尺度から予測することができます(Chapman1990)。

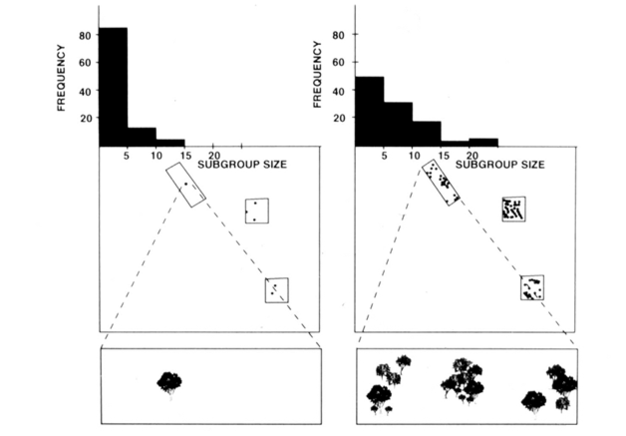
ウガンダのサンタローザ国立公園にあるクモザルのサブグループサイズが、毎月利用可能な食糧資源のサイズ、密度、分布に基づいて予測できることを この図の右側と左側は、異なる月を表しています。 左側には食べ物がまれな月です。 これは、正方形または長方形のサンプリングプロットと各プロット(最低パネル)の木を調べることによって見ることができます。 食べ物がまれな場合、大きなグループはパッチ(木)を急速に枯渇させ、グループが小さい場合に必要とされるよりも長い距離を移動する必要があります。 したがって、上のパネルに示されているように、食物が稀である場合、それが豊富であり、次のパッチまでの移動距離が小さい場合(図の右側)と比較して、サブグループはより小さくなる。
上記の種に加えて、生態学的制約モデルの適用性は、一般的に様々な種(Snaith&Chapman2005)および状況(例えば、general models、Wrangham et al. 1993年、および混合種協会、Chapman&Chapman2000c)。 我々は、このモデルが受けた広範な支持を考えると、種が典型的に生態学的制約モデルの期待に適合するべきであることを示唆することは合理的であ これは、すべての種が適合することを意味するものではなく、研究者はモデルの例外を検索することができるので、この分野での研究のための刺激的 適合しない種は、生態学的期待に反する社会戦略を採用しているため、適合しない種はそうすると思われます。 これは、様々な社会戦略の潜在的な重要性を評価するための”尺度”を提供する(すなわち、生態学的モデルの期待から逸脱するほど、社会的選択的圧力がより重要である)。 私たちは簡単に私たちの以前の研究から二つの例を提供してみましょう。
雌のチンパンジーは雌のクモサルよりも孤独であったが、両者は同様の核分裂融合社会組織を持っている。 さらに、生態学的条件に基づいて、サブグループ内の雄と亜成体のチンパンジーの数を正確に予測することはできましたが、これらのサブグループ内の雌のチンパンジーの数を予測することはできませんでした(Chapman et al. 1995). 資源が非常に豊富で、ほとんどすべての男性が一つの大きなサブグループにいたとしても、女性はグループに入ることはほとんどなく、より大きなサブグループにいることのコストが捕食者の回避などの利益を上回ることを示唆している。 これは、これらの女性の幼児が捕食によって最も脅かされている年齢/性別のクラスになるという観点から予想外です。 これらの観察を説明するための一つの検証可能な仮説は、これら二つの種の連合の性質がグループメンバーシップの利益に影響を与える可能性がある 証拠は、チンパンジーとは異なり、クモ猿の女性は、多くの場合、連合のメンバーが食料への排他的なアクセスを可能にするために動作する連合を形成す したがって、クモのサルは、特定の個体に加わると、他の個体を排除することによって食物へのアクセスを増やすことができますが、チンパンジーの雌 1995).
私たちは、潜在的にグループにいることのコストと利益に影響を与える社会的要因によって引き起こされる別の興味深い偏差を特定し、これは赤 guerezas、Colobus guereza)キバレ国立公園、ウガンダで。 赤いコロバスは、平均65個体(25-127個体、Snaith et al. 2008年)、guerezasは平均6.5人(4-11人、Harris&Chapman2007)の小グループに住んでいます。 この違いにもかかわらず、二つの種によって食べられる植物には多くの類似性があります。 生態学的条件が二つのコロビン種の間のグループサイズの違いの原因であった場合、彼らの食事が異なり、彼らはグループサイズが小さいので、ゲレサスの家の範囲で食物の木の密度が低くなることを期待するでしょう。 しかし、彼らの食事は非常に似ています;赤いコロバスと赤いコロバスグループの家の範囲内に完全にその家の範囲を持っていたゲレザグループの間の食2%、一方、赤いコロバスの二つの隣接するグループのために、食事はわずか37.3%によって重複した(Chapman et al. 2002). 女性のゲレザの生殖成功は、より大きなグループがテイクオーバーと乳児またはより多くの摂食競争の高い率を経験するため、中小のグループで最大化されているようです(folivorous Trachypithecus phayreiおよびSemnopithecus spのスクランブル競争の証拠を参照してください。 しかし、ゲレサスの証拠の欠如は、食品パッチを枯渇させる)。 私たちは、ゲレサスで男性のテイクオーバーと幼児殺害の数を目撃しています(Onderdonk2000)-Teichroebを参照してください&sicotte(2008)コロビンの幼児殺傷に関する理論 明確ではないのは、乳児の危険性が、複数の男性グループの形成を可能にする高密度の食物への明らかなアクセスを有する場合、ゲレザグループの大きさをどのように制限するかということである。 いくつかの種では、より大きな女性のグループは、男性を移住するためのターゲットの多くであり、この状況は、乳児率を増加させる可能性があります-例えば、Semnopithecus sp。 (Borries1997),Theropithecus gelada(Dunbar1984),Alouatta seniculus(Crockett&Janson2000),Presbytis thomasi(Steenbeek&van Schaik2001). しかし、新生児が将来の乳児のいくつかの可能性のある種牡馬に直面したときに、男性の移民とその後の暴力的な引き継ぎと乳児が発生する可能性が低いため、グループに複数の生殖活動性の男性が含まれている場合、乳児の発生頻度は低くなると予想される(Janson&van Schaik2000)。 Guerezasは、より大きなグループを形成するために十分な食料資源を持っているように見える(Tombak et al. 2012)、これは乳児のリスクを減少させるだろうが、まだ彼らは大きなグループを形成していません。 Guerezasにおける男性-男性相互作用の性質は、大きなグループを不安定にする可能性があります。 これはursine colobus(Colobus vellerosus)の場合のようであり、複数の男性グループが男性移民を引き付け、成人男性が一人の強いグループよりも幼児率が高い(Teichroeb,Wikberg,Badescu,&Sicotte、未公開データ;図3)。 Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

生態学的制約モデルが霊長類のグループサイズに一般的に影響を与える圧力を概説しており、この予想されるグループサイズからの逸脱が社会的圧力に起因する場合、それは興味深い質問の数を提起するので、この研究分野でエキサイティングな時間になるでしょう。 例えば: グループの規模に対する社会的圧力の性質は何ですか? これらの社会的制約は、これらの種に存在し、密接に関連していないのはなぜですか? そして、これらの社会的圧力のために進化の力は何を選択していますか? グループサイズは、種が採用する社会組織の種類に影響を与える重要な要素であり、これらの問題を調べることは、社会生態学的モデルの刺激的な精
Leave a Reply