Cytochemistryを祝う

繊毛および鞭毛は細胞からの突起である。 この漫画に示されているように、それらは微小管で構成されており、原形質膜の延長によって覆われています。 それらは運動性であり、細胞自体を移動させるか、または細胞の上または周囲の物質を移動させるように設計されている。 哺乳類細胞における繊毛の主な目的は、液体、粘液、または細胞をその表面上に移動させることである。 繊毛と鞭毛は同じ内部構造を持っています。 主な違いは、その長さです。

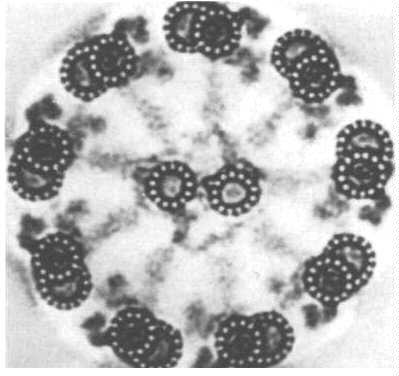
繊毛と鞭毛は、内部の一連の微小管の相互作用のために移動します。 これらを総称して”軸索”と呼び、この図は表面図と断面図(左下のパネル)に微小管(上部パネル)を示しています。 これらの微小管のうちの2つが結合して繊毛または鞭毛に1つの二重項を形成します。 尿細管の1つは不完全であることに注意してください。 さらに、微小管サブユニットの一つから突出する重要な微小管関連タンパク質(MAPs)がある。 この漫画では、繊毛の断面が示されています。
9つの二重小体の円があり、それぞれが1つの完全な(A細管)および1つの不完全な(B細管)微小管を有することに注意してください。 コアダブレットは両方とも完成しています。 Tから拡張する

彼のダブレットは、隣接するダブレットに参加するアームのセットです。 これらはタンパク質”dynein”で構成されています。 それは24nm間隔で間隔をあけられます。 Nexinリンクは、それらを一緒に保持するために微小管に沿って間隔をあけられている。 内側に突出しているのは、ダブルレットを囲むシースと接続する放射状のスポークです。
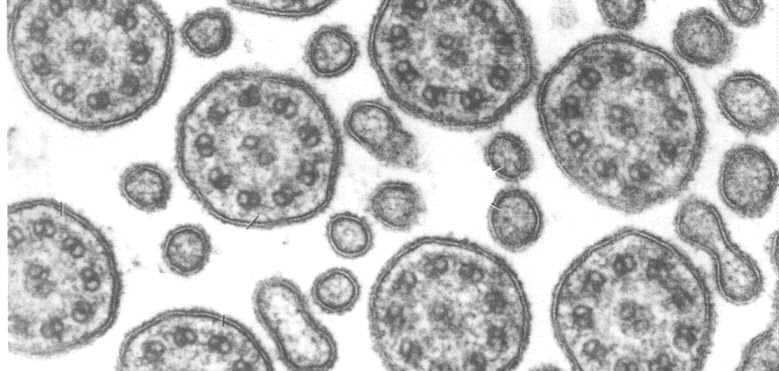
この図は、繊毛の断面の電子顕微鏡写真を示しています。 Dynein armsとnexinリンクが表示されることに注意してください。 Dyneinの腕にATPaseの活動があります。 ATPの存在下では、それらはあるチューブリンから別のチューブリンに移動することができる。 それらは細管が互いに沿って滑ることを可能にします従って繊毛は曲がることができます。
dynein橋は滑走が合わせられた曲がることをもたらすように調整される。 Nexinおよび放射状のスポークのために、doubletsは所定の位置に握られる従って滑走は縦に限られる。 Nexinおよび放射状のスポークが酵素の消化力に服従し、ATPに露出されれば、doubletは長さ9Xまで滑らせ、望遠鏡続けます。
以下は、繊毛の数を示す細胞表面の別の顕微鏡写真です。 これらは機能的に組織されなければならないので、繊毛は波の中で鼓動する。

繊毛と鞭毛は、細胞周辺に移動する中心小体から組織されています。 これらは”基底体”と呼ばれ、この電子顕微鏡写真(bb)に示されています。 細胞膜(cm)から突出した多数の繊毛に注意してください。 基底体は繊毛の動きの方向を制御する。 これは実験的に示すことができます。
中心小体は繊毛または鞭毛の方向を制御する運動。
Parameciumは、繊毛の平行な列をすべて揃えて、同じ方向に打つようにしています。 しかし、1960年代に繊毛/基底体の列がパラメシウムに移植され、ビートの方向に変化を示すことができた。 これは遺伝的変化ではなかったにもかかわらず、細胞は将来の世代に変化を伝えました。
セントリオール構造
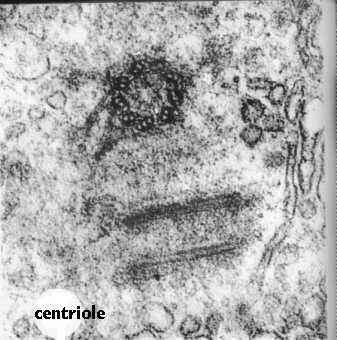
繊毛や鞭毛のように、セントリオールも微小管でできています。 違いは、それらが9組の三重項を含み、中央に二重項がないことです。 基底体の三重項がどのように繊毛二重項に変わるかは謎のままである。 中心小体は対になり、それぞれが他方に直角に組織される。 この図は、一対の中心小体の電子顕微鏡写真を示し、漫画は繊毛の断面を中心小体の断面と比較している。 中心小体は、有糸分裂の間に染色体が移動する紡錘体装置を組織する。

セントリオール複製

セントリオールは、ミトコンドリアやペルオキシソームのように自律的に複製します。 それらは、それらの形成に必要なタンパク質(チューブリンなど)を含む中心から始まる。)、その後、procentriolesフォーム。 それぞれは、三重項が形成することができる単一の微小管を成長させる。 中心小体が作られると、この漫画に示すように、娘の中心小体は尿細管から直角に成長することができます。 これらはその後、娘細胞(分裂細胞内)に追加されるか、またはそれらは周囲に移動し、繊毛の基底体を形成する。
詳細については、お問い合わせください:
グウェンチャイルズ、博士、FAAA
教授と椅子
神経生物学と発達科学の部門
アーカンソー大学医/div>質問については、このメールアドレスに連絡してください: http://www.cytochemistry.net
Leave a Reply