ce influențează dimensiunea grupurilor în care primatele aleg să trăiască?

una dintre contribuțiile teoretice majore aduse de studierea primatelor este o înțelegere a ceea ce influențează natura variației complexe în structura și organizațiile sociale. La baza acestui lucru se află un set de evoluții teoretice care examinează de ce animalele trăiesc în grupuri în primul rând și de ce grupurile sunt de o anumită dimensiune. Aici, ne concentrăm pe întrebarea ” Ce influențează dimensiunea grupurilor în care primatele aleg să trăiască?”
primatele sunt taxoni deosebit de utili pentru a aborda această întrebare, deoarece dimensiunile grupurilor lor sunt foarte variabile, nu numai în interiorul, ci și între specii. Mărimea grupului Intraspecific pentru colobus roșu (Procolobus rufomitratus), de exemplu, variază între 12 și 150 de membri (Chapman & Chapman 2000a). Variația interspecifică variază de la un singur individ, așa cum se vede la orangutani (Pongo sp.) (van Schaik 1999), la peste 800 de membri în mandrills (Mandrillus sphinx) (Abernethy și colab. 2002). Mai mult, în cadrul unor specii, grupurile sociale se împart în mod repetat și se reunesc în subgrupuri de diferite dimensiuni și combinații în timp (de exemplu, organizarea socială de fisiune-fuziune a maimuțelor păianjen (Ateles sp.) și cimpanzeii (Pan troglodytes), sau organizarea pe mai multe niveluri a gelada (Theropithecus gelada) și babuinii Hamadryas (Papio hamadryas, Aureli și colab. 2008). Această variație oferă fundamentul cercetătorilor pentru a dezvolta modele pentru a investiga atât factorii ecologici, cât și cei sociali ai dimensiunii grupului.
gruparea este benefică în mai multe moduri. Se crede că indivizii din grupuri mai mari au un risc scăzut de prădare (Hamilton 1971), pot fi mai capabili să găsească și să apere resursele alimentare (Cody 1971, Wrangham 1980) și pot fi protejați împotriva amenințărilor conspecifice, cum ar fi infanticidul de către bărbații din afara grupului (Wrangham 1979). Diversi cercetători au sugerat că gruparea conferă beneficii atât de previzibile (Alexander 1974, van Schaik 1983) încât diferențele de dimensiune a grupului pot fi explicate prin dezavantaje (Wrangham și colab. 1993). Cel mai larg acceptat cost potențial al grupării este considerat a fi o reducere a eficienței hrănirii. A fi cu alți indivizi cu aceleași cerințe dietetice înseamnă că animalele fie se luptă pentru hrană (concurs de concurs), fie un animal dintr-un grup bate altul la mâncare, astfel când al doilea animal vine într-o zonă, pur și simplu nu mai rămâne mâncare (scramble competition, Janson & van Schaik 1988). În ambele situații se crede că concurența asupra alimentelor duce la animale care trebuie să călătorească mai departe. Logica din spatele acestui argument este relativ simplă. Animalele trebuie să se hrănească într-o zonă care să le poată satisface cerințele energetice și nutriționale. Rezultă că o creștere a dimensiunii grupului va crește suprafața care trebuie acoperită pentru a găsi provizii alimentare adecvate. Astfel, indivizii trebuie să călătorească mai departe și să cheltuiască mai multă energie dacă se află într-un grup mare, decât dacă se hrănesc într-un grup mai mic. Odată cu creșterea timpului petrecut în călătorie, se apropie un punct în care energia cheltuită în călătorie este prea costisitoare și grupurile mai mici devin avantajoase. În acest fel, factorii ecologici pot influența modelele de mișcare și eficiența hrănirii, constrângând astfel dimensiunea grupurilor care pot exploata eficient resursele alimentare disponibile. Aceste idei au fost formalizate în ceea ce a devenit cunoscut sub numele de modelul constrângerilor ecologice (Chapman & Chapman 2000B, Ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
componenta esențială a modelului constrângerilor ecologice este că o creștere a dimensiunii grupului trebuie să conducă la o creștere a concurenței de hrănire în cadrul grupului. Este posibil ca aceasta să funcționeze într-un mod ușor diferit, în funcție de natura resurselor utilizate de anumite specii. Cu primate frugivore și, eventual, multe folivore, care se hrănesc în pete discrete — de obicei copaci care poartă produse alimentare sau aglomerări de copaci — membrii suplimentari ai grupului pot epuiza petele mai repede și pot duce la intervale de zi crescute (Chapman 1988, Snaith & Chapman 2005). Pentru speciile mai insectivore, ale căror resurse pot să nu apară în patch-uri discrete, călătoria continuă pe baldachin în căutarea insectelor cu membri suplimentari ai grupului poate duce la o creștere a suprapunerii câmpurilor de căutare individuale, reducând ratele de întâlnire pe cap de locuitor cu alimentele și crescând astfel zona care trebuie căutată (van Schaik și colab. 1983).
deci, modelul constrângerilor ecologice sugerează că acei factori care afectează distanța pe care o parcurg animalele ar trebui să afecteze și dimensiunea grupului. Pentru acele animale care se hrănesc de obicei cu fructe sau frunze care pot fi epuizate, dimensiunea plasturelui ar determina cât timp ar putea rămâne și hrăni un grup de o anumită dimensiune. Un grup mare ar petrece mai puțin timp într — un plasture de o anumită dimensiune decât un grup mai mic, deoarece epuizează plasturele mai repede-un grup mare are pur și simplu mai multe guri de hrănit. Dacă animalele călătoresc între patch-uri după ce le-au epuizat, atunci densitatea și distribuția patch-urilor vor determina costurile de călătorie suportate. Atunci când patch-urile de resurse sunt la o densitate mare sau într-o distribuție aglomerată, Distanța până la următorul patch este mică, costurile de călătorie sunt scăzute și, prin urmare, animalele pot forma grupuri mari. În astfel de momente, orice cost suplimentar asociat cu a fi membru al unui grup mare, cum ar fi necesitatea de a vizita multe patch-uri, poate fi ușor recuperat. În schimb, atunci când patch-urile de resurse apar la densități mici, Distanța până la următorul patch este de obicei mare, costurile de călătorie sunt mari, iar animalele nu își pot permite să epuizeze rapid patch-urile și, prin urmare, animalele formează grupuri mici.
organizarea socială de fisiune-fuziune a maimuțelor păianjen, a cimpanzeilor și a altor câteva primate (Figura 1) oferă teste utile ale modelului de constrângeri ecologice, deoarece se poate încerca prezicerea schimbărilor temporale ale dimensiunii subgrupului din măsurile directe ale dimensiunii, densității și distribuției resurselor alimentare (Chapman 1990, Chapman și colab. 1995). Pentru maimuțele păianjen și cimpanzeii, a fost dezvoltat un model de regresie multiplă pentru a arăta că dimensiunea, densitatea și distribuția petelor alimentare măsurate în fiecare lună au fost un bun predictor al dimensiunii subgrupului. Este dificil să se reprezinte vizual o regresie multiplă, dar metodele utilizate pentru maimuțele păianjen sunt prezentate în Figura 2. Maimuțele păianjen au un tip de organizare socială foarte flexibil de fisiune-fuziune, iar în Parcul Național Santa Rosa, Costa Rica, dimensiunea subgrupului poate varia de la 1 la 35 de persoane, dar în medie se găsesc aproximativ 5 persoane care călătoresc împreună. Pentru această populație, 50% din varianța dimensiunii medii lunare a subgrupului poate fi prezisă din măsuri relativ brute ale dimensiunii, densității și distribuției patch-urilor alimentare (Chapman 1990).

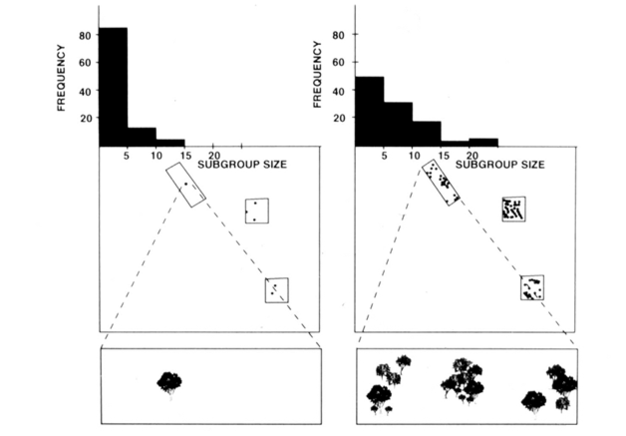
În plus față de speciile menționate mai sus, aplicabilitatea modelului constrângerilor ecologice a fost în general susținută cu cercetări provenind dintr-o varietate de specii (Snaith & Chapman 2005) și situații (de exemplu, modele generale, Wrangham și colab. 1993, și asociații de specii mixte, Chapman & Chapman 2000C). Considerăm că, având în vedere sprijinul larg pe care l-a primit acest model, a sosit momentul să se sugereze că speciile ar trebui să se conformeze în mod obișnuit așteptărilor modelului de constrângeri ecologice. Acest lucru nu înseamnă că toate speciile se vor conforma și, astfel, devine un moment interesant pentru cercetarea în acest domeniu, deoarece anchetatorii pot căuta excepții de la model. Bănuim că speciile care nu se conformează vor face acest lucru deoarece au adoptat strategii sociale care contravin așteptărilor ecologice. Aceasta oferă un „etalon” pentru a evalua importanța potențială a diferitelor strategii sociale (adică, cu cât se abate în continuare de la așteptările modelului ecologic, cu atât sunt mai importante presiunile selective sociale).
Să oferim pe scurt două exemple din cercetările noastre anterioare. Cimpanzeii de sex feminin erau mai solitari decât maimuțele păianjen de sex feminin, totuși ambele au organizații sociale similare de fisiune-fuziune. Mai mult, în timp ce am putea prezice cu exactitate numărul de bărbați și cimpanzei subadulți într-un subgrup bazat pe condiții ecologice, nu am putut prezice numărul de cimpanzei femele din aceste subgrupuri (Chapman și colab. 1995). Chiar și atunci când resursele erau extrem de abundente și aproape toți bărbații se aflau într-un subgrup mare, femelele au intrat rar în grupuri, sugerând că costul de a fi într-un subgrup mai mare a depășit orice beneficii, cum ar fi evitarea prădătorilor. Acest lucru este neașteptat din perspectiva că sugarii acestor femele ar fi clasa de vârstă/sex cea mai amenințată de prădare. O ipoteză testabilă pentru a explica aceste observații este că natura coalițiilor din aceste două specii poate influența beneficiile apartenenței la grup. Dovezile sugerează că, spre deosebire de cimpanzei, femelele maimuță păianjen formează coaliții care funcționează adesea pentru a permite membrilor coaliției acces exclusiv la hrană. Așadar, maimuțele păianjen, atunci când se alătură anumitor indivizi, își pot crește accesul la hrană prin excluderea altora, dar pentru femelele de cimpanzeu acest lucru nu este cazul (Chapman și colab. 1995).
am identificat o altă abatere interesantă, potențial cauzată de factori sociali care afectează costurile și beneficiile de a fi într-un grup, iar aceasta se referă la Colobus roșu și maimuțe colobus alb-negru (aka. guerezas, Colobus guereza) în Parcul Național Kibale, Uganda. Colobusul roșu formează grupuri mari cu o medie de 65 de indivizi (25-127 indivizi, Snaith și colab. 2008), în timp ce guerezas trăiesc în grupuri mici, cu o medie de 6,5 indivizi (4-11 indivizi, Harris & Chapman 2007). În ciuda acestei diferențe, există o mare asemănare în plantele consumate de cele două specii. Dacă condițiile ecologice ar fi responsabile pentru diferența de dimensiune a grupului între cele două specii de colobine, s-ar aștepta ca dietele lor să difere și densitatea copacilor alimentari să fie mai mică în intervalele de origine ale guerezelor, deoarece acestea au dimensiunea grupului mai mică. Cu toate acestea, dietele lor sunt foarte asemănătoare; suprapunerea dietetică între un colobus roșu și un grup guereza care și-a avut gama de origine în întregime în gama de origine a grupului Colobus roșu a fost de 43.2%, în timp ce pentru cele două grupuri vecine de colobus roșu, dieta s-a suprapus cu doar 37,3% (Chapman și colab. 2002). Se pare că succesul reproductiv al femeilor guereza este maximizat în grupurile mici și mijlocii, fie pentru că grupurile mai mari se confruntă cu rate mai mari de preluări și infanticid, fie cu o concurență mai mare de hrănire (vezi dovezi ale concurenței de luptă în folivor Trachypithecus phayrei și Semnopithecus sp. , dar lipsa dovezilor că guerezas epuizează patch-urile alimentare ). Am asistat la o serie de preluări de bărbați și ucideri de copii în guerezas (Onderdonk 2000)-vezi Teichroeb & Sicotte (2008) pentru o evaluare a teoriei privind infanticidul la colobine. Ceea ce nu este clar este modul în care riscul de infanticid ar limita dimensiunea grupurilor de guereza, atunci când acestea au acces aparent la o densitate mare de alimente care ar trebui să permită formarea grupurilor multi-masculine. La mai multe specii, grupurile mai mari de femei sunt mai mult o țintă pentru bărbații care emigrează, iar această situație poate crește ratele de infanticid-de exemplu, Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). Cu toate acestea, infanticidul este de așteptat să fie mai puțin frecvent dacă un grup conține mai mulți bărbați activi reproductiv, deoarece imigrația masculină și preluarea violentă ulterioară și infanticidul sunt mai puțin susceptibile să apară atunci când noul venit se confruntă cu mai multe siruri posibile ale viitorilor sugari (Janson & van Schaik 2000). Guerezele par să aibă resurse alimentare ample pentru a forma grupuri mai mari (Tombak și colab. 2012), ceea ce ar reduce riscul de infanticid, dar nu formează grupuri mari. Este posibil ca natura interacțiunilor Bărbat-Bărbat În guerezas să facă grupuri mari instabile. Acest lucru pare să fie cazul în ursine colobus( Colobus vellerosus), unde grupurile multi-masculine atrag imigranți de sex masculin și au rate mai mari de infanticid decât grupurile cu un bărbat adult puternic în vârstă (Teichroeb, Wikberg, Badescu, & Sicotte, date nepublicate; Figura 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

dacă modelul constrângerilor ecologice conturează presiunile care influențează în general dimensiunea grupului primatelor și dacă abaterile de la această dimensiune așteptată a grupului rezultă din presiunile sociale, va fi un moment interesant în acest domeniu de cercetare, deoarece ridică o serie de întrebări interesante. De exemplu: Care este natura presiunilor sociale asupra dimensiunii grupului? De ce sunt prezente aceste constrângeri sociale la aceste specii și nu sunt strâns legate? Și ce aleg forțele evolutive pentru aceste presiuni sociale? Dimensiunea grupului este un element critic care influențează tipul de organizare socială pe care o adoptă speciile, examinând astfel aceste probleme va duce la rafinări interesante ale modelelor socioecologice.
Leave a Reply