8. 5: Reseptorityrosiinikinaasit (RTKs)
Reseptorityrosiinikinaasit välittävät vasteita suurelle joukolle signaaleja, mukaan lukien peptidihormonit kuten insuliini ja kasvutekijät kuten epidermaalinen kasvutekijä. Kuten GPC-reseptorit, reseptorityrosiinikinaasit sitovat signaalin ja välittävät viestin solunsisäisten molekyylien läpi, joista viimeinen vaikuttaa kohdeproteiineihin solun tilan muuttamiseksi.
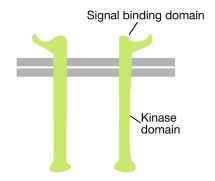
kuten nimestä voi päätellä, reseptorityrosiinikinaasi on solun pintareseptori, jolla on myös tyrosiinikinaasiaktiivisuus. Reseptorityrosiinikinaasin signaalia sitova domeeni on solun pinnalla, kun taas tyrosiinikinaasin entsymaattinen aktiivisuus sijaitsee proteiinin sytoplasmaosassa (KS.kuva yllä). Transmembraani alfa helix yhdistää nämä kaksi aluetta reseptorin.
Mitä tapahtuu, kun signaalimolekyylit sitoutuvat reseptorityrosiinikinaaseihin?
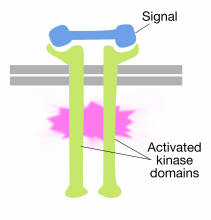
signaalimolekyylien sitoutuminen reseptorin solunulkoisiin domeeneihin tyrosiinikinaasimolekyylit aiheuttavat kahden reseptorimolekyylin dimerisoitumisen (tulevat yhteen ja liittyvät). Tämä tuo reseptorien sytoplasmahännät lähelle toisiaan ja saa näiden häntien tyrosiinikinaasiaktiivisuuden käynnistymään. Tämän jälkeen aktivoidut hännät fosforyloivat toisiaan useille tyrosiinijäämille. Tätä kutsutaan autofosforylaatioksi.
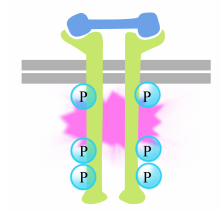
tyrosiinien fosforylaatio reseptorihännissä laukaisee solunsisäinen signalointikompleksi hännässä. Vasta fosforyloidut tyrosiinit toimivat sitoutumispaikkoina proteiineille, jotka sitten välittävät viestin edelleen muille proteiineille. Tärkeää proteiinia, jota reseptorityrosiinikinaasien signalointikompleksit aktivoivat, kutsutaan Ras: ksi.
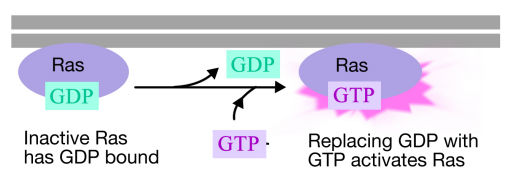
Ras-proteiini on monomeerinen guaniininukleotidia sitova proteiini, joka liittyy plasman solukalvon sytosolipintaan (itse asiassa se muistuttaa paljon trimeeristen G-proteiinien alfa-alayksikköä). Aivan kuten G – proteiinin alfa-alayksikkö, Ras on aktiivinen, kun GTP on sitoutunut siihen, ja inaktiivinen, kun BKT on sitoutunut siihen.Myös, kuten alfa-alayksikkö, Ras voi hydrolysoida GTP: n BKT: ksi.
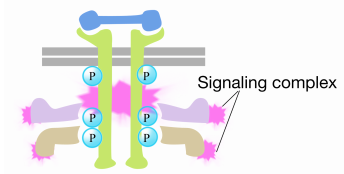
kun signaali saapuu reseptorityrosiinikinaasiin, reseptorimonomeerit yhtyvät ja fosforyloivat toistensa tyrosiineja, mikä laukaisee proteiinikompleksin kokoontumisen reseptorin sytoplasmaiseen häntään. Yksi tämän kompleksin proteiineista vuorovaikuttaa Ras: n kanssa ja stimuloi inaktiiviseen Ras: ään sitoutuneen BKT: n vaihtoa GTP: hen. Tämä aktivoi Ras: n.
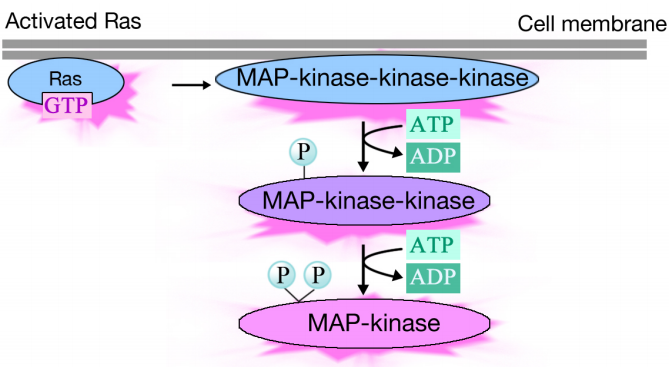
aktivoitu Ras laukaisee kolmen proteiinikinaasin fosforylaatiokaskadin, joka välittää ja jakaa signaalin. Nämä proteiinikinaasit kuuluvat ryhmään nimeltä MAP-kinaasit (Mitogeenin Aktivoimat Proteiinikinaasit). Lopullinen kinaasi tässä kaskadissa fosforyloi erilaisia kohdeproteiineja, kuten entsyymejä ja transkriptioaktivaattoreita, jotka säätelevät geenien ilmentymistä.
eri entsyymien fosforylaatio voi muuttaa niiden toimintaa ja käynnistää uusia kemiallisia reaktioita solussa, kun taas transkriptioaktivaattoreiden fosforylaatio voi muuttaa, mitkä geenit ilmentyvät. Geeniekspression ja proteiinin aktiivisuuden muutosten yhteisvaikutus muuttaa solun fysiologista tilaa.
jälleen kerran seuratessa RTKs: n välittämää signaalinvälityspolkua on mahdollista havaita sama tapahtumien peruskuvio: signaali sitoutuu reseptorityrosiinikinaasien solunulkoisiin domeeneihin, jolloin sytosolihännät dimerisoituvat ja autofosforyloituvat, jolloin viesti välittyy solun sisäosiin.
viesti välitetään Merkinantokompleksin kautta Ras: lle, joka sitten stimuloi kinaasisarjaa. Kaskadissa oleva terminaalinen kinaasi vaikuttaa kohdeproteiineihin ja saa aikaan muutoksia proteiinin toiminnassa ja geeniekspressiossa.
yllä olevat kuvaukset tarjoavat hyvin yksinkertaisen luonnoksen joistakin reseptoriluokista ja käsittelevät pääasiassa mekanistisia yksityiskohtia vaiheista, joilla erityyppisten reseptoreiden vastaanottamat signaalit saavat aikaan muutoksia soluissa. Yksi tärkeä kotiopetus on eri reittien olennainen samankaltaisuus.
toinen muistettava seikka on se, että kun olemme tarkastelleet jokaista yksittäistä reittiä eristyksissä, solu saa milloin tahansa useita signaaleja, jotka laukaisevat useita erilaisia vasteita kerralla. Edellä kuvatut reitit osoittavat huomattavaa ”ristipuhetta”, ja minkä tahansa signaalin vasteeseen vaikuttavat solun samanaikaisesti vastaanottamat muut signaalit. Erilaisten reseptorien, signaalien ja niiden yhdistelmien moninaisuus on keino, jolla solut pystyvät reagoimaan valtavaan moninaisuuteen erilaisissa olosuhteissa.
avustajat
-
tohtori Kevin Ahern ja tohtori Indira Rajagopal (Oregon State University)
Leave a Reply