What Influences the Size of Groups in Which kädelliset Choose to Live?

yksi kädellisten tutkimisen keskeisistä teoreettisista panoksista on ymmärrys siitä, mikä vaikuttaa yhteiskuntarakenteen ja organisaatioiden monimutkaisen vaihtelun luonteeseen. Taustalla on joukko teoreettisia kehityskulkuja, joissa tutkitaan, miksi eläimet ylipäätään elävät ryhmissä ja miksi ryhmät ovat tietyn kokoisia. Tässä keskitymme kysymykseen ” mikä vaikuttaa ryhmien kokoon kädelliset haluavat elää?”
kädelliset ovat erityisen hyödyllisiä taksoneita tämän kysymyksen ratkaisemiseksi, koska niiden ryhmäkoot vaihtelevat suuresti paitsi lajien sisällä myös niiden välillä. Esimerkiksi punakolobuksen (”Procolobus rufomitratus”) ryhmäkoko vaihtelee 12-150 jäsenen välillä (Chapman & Chapman 2000a). Lajienvälinen vaihtelu vaihtelee yksittäisestä yksilöstä, kuten orangeilla (Pongo sp.) (van Schaik 1999), yli 800: lle mandrillien (Mandrillus sphinx) jäsenelle (Abernethy et al. 2002). Lisäksi joidenkin lajien sisällä sosiaaliset ryhmät jakautuvat toistuvasti ja yhdistyvät uudelleen eri kokoisiksi ja eri yhdistelmiksi ajan kuluessa (esim. hämähäkkiapinoiden fissiofuusiososiaalinen organisaatio (Ateles sp.) ja simpanssit (Pan troglodytes) eli geladan (Theropithecus gelada) ja Hamadryas-paviaanien (Papio hamadryas, Aureli et al. 2008). Tämä muunnelma antaa tutkijoille pohjan kehittää malleja, joilla tutkitaan sekä ryhmäkokoon liittyviä ekologisia että sosiaalisia tekijöitä.
ryhmittelystä on hyötyä monella tavalla. Suurempien ryhmien yksilöillä arvellaan olevan vähentynyt saalistusriski (Hamilton 1971), ne saattavat pystyä paremmin löytämään ja puolustamaan ravintovaroja (Cody 1971, Wrangham 1980) ja niitä voidaan suojella erityiseltä uhalta, kuten ryhmän ulkopuolisten urosten lapsenmurhalta (Wrangham 1979). Eri tutkijat ovat ehdottaneet, että ryhmittely antaa sellaisia ennustettavia etuja (Alexander 1974, van Schaik 1983), että ryhmäkokojen erot voidaan selittää haitoilla (Wrangham et al. 1993). Laajimmin hyväksyttyjen ryhmittelykustannusten arvellaan vähentävän ravinnonhankinnan tehokkuutta. Muiden samojen ravintovaatimusten omaavien yksilöiden kanssa oleminen tarkoittaa sitä, että eläimet joko taistelevat ruoasta (kilpailu) tai yksi eläin ryhmässä voittaa toisen ravinnosta, jolloin toisen eläimen tullessa alueelle ruokaa ei yksinkertaisesti ole jäljellä (scramble competition, Janson & van Schaik 1988). Molemmissa tilanteissa ajatellaan, että kilpailu ravinnosta johtaa siihen, että eläimet joutuvat matkustamaan kauemmas. Tämän väitteen logiikka on suhteellisen yksinkertainen. Eläinten on syötävä alueella, joka täyttää niiden energia-ja ravintotarpeet. Tästä seuraa, että ryhmäkokojen kasvattaminen lisää alaa, joka on katettava riittävän ravinnon saamiseksi. Näin ollen yksilöiden täytyy matkustaa kauemmas ja kuluttaa enemmän energiaa, jos he ovat suuressa ryhmässä, kuin jos he syövät pienemmässä ryhmässä. Kun matkustamiseen käytetty aika kasvaa, lähestytään kohtaa, jossa matkustamiseen käytetty energia on liian kallista ja pienemmistä ryhmistä tulee edullisia. Tällä tavoin ekologiset tekijät voivat vaikuttaa liikkumistapoihin ja ravinnon etsinnän tehokkuuteen, mikä rajoittaa sellaisten ryhmien kokoa, jotka voivat tehokkaasti hyödyntää saatavilla olevia ravintoresursseja. Nämä ajatukset on virallistettu niin sanotussa ekologisten rajoitteiden mallissa (Chapman & Chapman 2000B, Ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
ekologisten rajoitteiden mallin olennainen osa on se, että ryhmäkoon kasvattamisen on johdettava ryhmän sisäisen ruokintakilpailun lisääntymiseen. On mahdollista, että tämä toimii hieman eri tavalla riippuen tietyn lajin käyttämien resurssien luonteesta. Frugivorous, ja mahdollisesti monia folivorous kädellisiä, jotka syövät erillisiä laikkuja-tyypillisesti puita, joissa on elintarvikkeita tai puun möhkäleitä-lisää ryhmän jäseniä voi poistaa laikkuja nopeammin ja johtaa lisääntyneisiin päiviin (Chapman 1988, Snaith & Chapman 2005). Enemmän hyönteissyöjälajeja, joiden luonnonvarat eivät välttämättä esiinny erillisinä laikkuina, jatkuva matkustaminen katoksen läpi hyönteisten etsimisessä ylimääräisten ryhmän jäsenten kanssa voi johtaa yksittäisten hakukenttien päällekkäisyyksien lisääntymiseen, vähentäen henkeä kohti kohtaamisasteita ruoan kanssa ja siten lisäämällä etsittävää aluetta (van Schaik et al. 1983).
ekologisten rajoitteiden mallin mukaan niiden tekijöiden, jotka vaikuttavat eläinten kulkemaan etäisyyteen, pitäisi vaikuttaa myös ryhmäkokoon. Niiden eläinten osalta, jotka yleensä käyttävät ravinnokseen hedelmiä tai lehtiä, jotka voivat ehtyä, laastarin koko määrittäisi, kuinka kauan tietyn kokoinen ryhmä voi viipyä ja ruokkia. Suuri ryhmä viettäisi vähemmän aikaa tietyn kokoisessa laastarissa kuin pienempi ryhmä, koska se kuluttaa laastarin nopeammin — suurella ryhmällä on yksinkertaisesti enemmän suuta ruokittavana. Jos eläimet liikkuvat laikkujen välillä, kun ne ovat ehtyneet, niin laikkujen tiheys ja jakautuminen määräävät matkakustannukset. Kun resurssilaastarit ovat tiheässä tai jakaantuneina, etäisyys seuraavaan paikkaan on pieni, matkakustannukset ovat pienet ja eläimet voivat siksi muodostaa suuria ryhmiä. Tällaisina aikoina kaikki suuren ryhmän jäsenyyteen liittyvät lisäkustannukset, kuten tarve käydä monissa laastareissa, voidaan helposti periä takaisin. Kun taas resurssilaastareita esiintyy harvoissa tiheyksissä, etäisyys seuraavaan laastariin on tyypillisesti suuri, matkakustannukset ovat korkeat, eikä eläimillä ole varaa hävittää laastareita nopeasti, minkä vuoksi eläimet muodostavat pieniä ryhmiä.
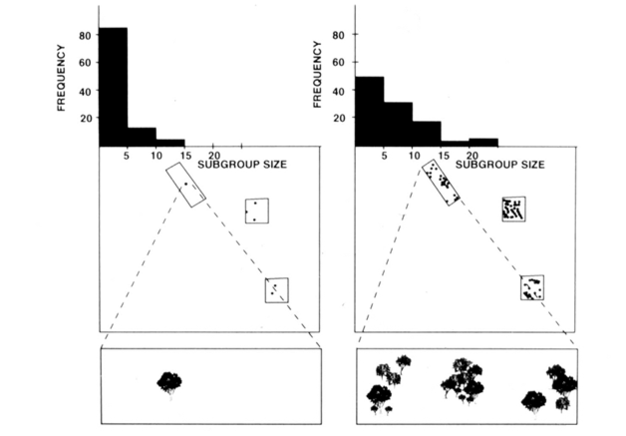
hämähäkkiapinoiden, simpanssien ja muutamien muiden kädellisten fissiofuusiososiaalinen organisaatio (Kuva 1) tarjoaa hyödyllisiä ekologisten rajoitteiden mallin testejä, koska alaryhmien koon ajallisia muutoksia voidaan yrittää ennustaa suoraan mittaamalla ruokavarojen kokoa, tiheyttä ja jakautumista (Chapman 1990, Chapman et al. 1995). Hämähäkkiapinoille ja simpansseille kehitettiin moninkertainen regressiomalli, joka osoitti, että kuukausittain mitattujen ruokalappujen koko, tiheys ja jakautuminen olivat hyvä alaryhmien koon ennustaja. Moninkertainen regressio on vaikea esittää visuaalisesti, mutta hämähäkkiapinoille käytetyt menetelmät esitetään kuvassa 2. Hämähäkkiapinoilla on hyvin joustava fissiofuusiotyyppinen sosiaalinen organisaatio, ja Santa Rosan kansallispuistossa Costa Ricassa aliryhmän koko voi vaihdella 1-35 yksilöstä, mutta keskimäärin noin 5 yksilöä tavataan matkustamassa yhdessä. Tälle populaatiolle 50% kuukausittaisen keskikoonvaihtelusta voidaan ennustaa elintarvikelaastareiden koon, tiheyden ja jakauman suhteellisen karkeista mittauksista (Chapman 1990).

edellä mainittujen lajien lisäksi ekologisten rajoitteiden mallin sovellettavuutta on yleisesti tuettu eri lajeista (Snaith& Chapman 2005) ja tilanteista (esim.general models, Wrangham et al. 1993, ja lajiseurat, Chapman & Chapman 2000C). Kun otetaan huomioon tämän mallin saama laaja tuki, on mielestämme tullut aika, jolloin on järkevää ehdottaa, että lajien olisi yleensä vastattava ekologisten rajoitusten mallin odotuksia. Tämä ei tarkoita sitä, että kaikki lajit mukautuisivat, ja siten siitä tulee jännittävä aika tämän alan tutkimukselle, koska tutkijat voivat etsiä poikkeuksia malliin. Epäilemme, että lajit, jotka eivät noudata vaatimuksia, tekevät niin, koska ne ovat omaksuneet sosiaalisia strategioita, jotka ovat ristiriidassa ekologisten odotusten kanssa. Tämä tarjoaa ”mittapuun” erilaisten sosiaalisten strategioiden mahdollisen merkityksen arvioimiseksi (ts.mitä pidemmälle ekologisen mallin odotuksista poiketaan, sitä tärkeämpiä ovat sosiaaliset valikoivat paineet).
esittäkäämme lyhyesti kaksi esimerkkiä aiemmasta tutkimuksestamme. Naarassimpanssit olivat yksinäisempiä kuin naarashämähäkkiapinat, mutta molemmilla on samanlaiset fissiofuusiososiaaliset organisaatiot. Edelleen, vaikka pystyimme ennustamaan tarkasti urosten ja alempien simpanssien määrän alaryhmässä ekologisten olosuhteiden perusteella, emme pystyneet ennustamaan naarassimpanssien määrää näissä alaryhmissä(Chapman et al. 1995). Silloinkin kun luonnonvarat olivat erittäin runsaat ja lähes kaikki urokset olivat yhdessä suuressa alaryhmässä, naaraat siirtyivät harvoin ryhmiin, mikä viittaa siihen, että suuremmassa alaryhmässä olemisen kustannukset olivat suuremmat kuin mahdolliset hyödyt, kuten saalistajien välttäminen. Tämä on odottamatonta siitä näkökulmasta, että näiden naaraiden poikaset olisivat se Ikä – /sukupuoliluokka, jota saalistusuhka eniten uhkaa. Yksi testattavissa oleva hypoteesi näiden havaintojen selittämiseksi on, että näiden kahden lajin yhteenliittymien luonne voi vaikuttaa ryhmän jäsenyyden hyötyihin. Todisteet viittaavat siihen, että toisin kuin simpanssit, hämähäkkiapinanaaraat muodostavat koalitioita, jotka usein toimivat salliakseen koalition jäsenille yksinoikeuden saada ruokaa. Joten hämähäkkiapinat, kun ne liittyvät tiettyihin yksilöihin, voivat lisätä ruoan saantia sulkemalla muut pois, mutta simpanssinaaraiden kohdalla näin ei ole (Chapman et al. 1995).
tunnistimme toisen mielenkiintoisen poikkeaman, joka saattaa johtua sosiaalisista tekijöistä, jotka vaikuttavat ryhmään kuulumisen kustannuksiin ja hyötyihin, ja tämä koskee punakolobusapinoita ja mustavalkoisia kolobusapinoita (aka. guerezas, Colobus guereza) kibalen kansallispuistossa Ugandassa. Punakolobut muodostavat suuria ryhmiä, joissa on keskimäärin 65 yksilöä(25-127 yksilöä, Snaith ym. 2008), kun taas guerezat elävät pienissä ryhmissä keskimäärin 6,5 yksilöä (4-11 yksilöä, Harris & Chapman 2007). Tästä erosta huolimatta näiden kahden lajin syömissä kasveissa on paljon samankaltaisuutta. Jos ekologiset olosuhteet olisivat syynä kahden kolobiinilajin ryhmäkoon eroon, voisi olettaa, että niiden ruokavalio olisi erilainen ja ravintopuiden tiheys olisi pienempi guerezojen kotialueilla, koska niillä on pienempi ryhmäkoko. Niiden ruokavalio on kuitenkin hyvin samankaltainen; punakolobus-ryhmän ja guereza-ryhmän, jonka kotoperäinen levinneisyysalue oli kokonaan punakolobus-ryhmän kotoperäinen, ravinnon päällekkäisyys oli 43.2%, kun taas kahden naapuriryhmän punakolobus, ruokavalio päällekkäin vain 37,3% (Chapman et al. 2002). Näyttää siltä, että naaras guereza lisääntymismenestys on maksimoitu pienissä ja keskisuurissa ryhmissä, joko siksi, että suuremmilla ryhmillä on enemmän ottoja ja lapsenmurhia tai enemmän ruokintakilpailua (KS.todisteita sekoituskilpailusta folivorous Trachypithecus phayrei ja Semnopithecus sp. , mutta ei ole näyttöä guerezas heikentävät elintarvikelaastareita). Guerezoissa on nähty useita miesten valtauksia ja lapsenmurhia (Onderdonk 2000)-katso Teichroeb & Sicotte (2008) kolobiinien lapsenmurhia koskevan teorian arvioinnista. Ei ole selvää, miten lapsenmurhien riski rajoittaisi guereza-ryhmien kokoa, kun niillä on nähtävästi käytettävissään runsaasti ravintoa, jonka pitäisi mahdollistaa usean miehen ryhmien muodostuminen. Useissa lajeissa isommat naarasryhmät ovat enemmänkin kohde muuttaville uroksille, ja tämä tilanne voi lisätä lapsenmurhien määrää-esimerkiksi Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). Lapsenmurhien odotetaan kuitenkin olevan harvinaisempia, jos ryhmässä on useita lisääntymiskykyisiä uroksia, koska miesten maahanmuutto ja sitä seuraava väkivaltainen valtaus ja lapsenmurhat ovat epätodennäköisempiä, kun tulokas kohtaa useita mahdollisia tulevien imeväisten poikasia (Janson & van Schaik 2000). Guerezat näyttävät olevan runsaasti ruokaa resursseja muodostaa suurempia ryhmiä (Tombak et al. 2012), mikä vähentäisi lapsenmurhien riskiä, mutta ne eivät kuitenkaan muodosta suuria ryhmiä. On mahdollista, että guerezoilla esiintyvien miesten ja miesten välisten vuorovaikutusten luonne tekee suurista ryhmistä epävakaita. Näin näyttää olevan ursine colobuksessa (Colobus vellerosus), jossa monen miehen ryhmät vetävät puoleensa miespuolisia maahanmuuttajia ja joissa on enemmän lapsenmurhia kuin ryhmissä, joissa on yksi vahva aikuinen mies parhaassa iässä (Teichroeb, Wikberg, Badescu, & Sicotte, julkaisemattomat tiedot; kuva 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

Esimerkiksi: Millaisia ovat ryhmäkokoon liittyvät sosiaaliset paineet? Miksi nämä sosiaaliset rajoitteet ovat läsnä näissä lajeissa eivätkä läheistä sukua? Ja mitä evolutionaariset voimat valitsevat näihin yhteiskunnallisiin paineisiin? Ryhmäkoko on kriittinen tekijä, joka vaikuttaa siihen, millaisen sosiaalisen organisaation lajit omaksuvat, joten näiden kysymysten tarkastelu johtaa jännittäviin sosioekologisten mallien tarkennuksiin.
Leave a Reply