8.5: Receptor Tirozin Kinázok (RTKs)
Receptor tirozin kinázok mediációt a válaszok nagy jelek száma, beleértve a peptid hormonok, mint az inzulin, valamint a növekedési tényezők, mint epidermális növekedési faktor. Mint a határok között mozog, receptor tirozin kinázok bind egy jelet, akkor adja át az üzenetet egy sor intracelluláris molekulák, amelyek közül az utolsó hat cél fehérjék változás, hogy az állam a sejt.
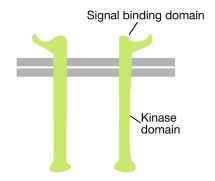
ahogy a neve is sugallja, a receptor tirozin-kináz olyan sejtfelszíni receptor, amely tirozin-kináz aktivitással is rendelkezik. A receptor tirozin-kináz jelkötő doménje a sejt felületén található, míg a tirozin-kináz enzimaktivitás a fehérje citoplazmatikus részében található (lásd a fenti ábrát). A transzmembrán alfa hélix összeköti a receptor e két régióját.
mi történik, ha a jelmolekulák kötődnek a receptor tirozin kinázokhoz?
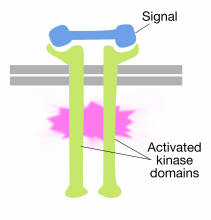
Kötés jel molekulák a sejten kívüli domain a receptor tirozin kináz molekulák okoz a két receptor molekulák dimerize (össze munkatárs). Ez a receptorok citoplazmatikus farkait egymáshoz közel hozza, és ezeknek a faroknak a tirozin-kináz aktivitását bekapcsolja. Az aktivált farok ezután foszforilálják egymást több tirozin maradékon. Ezt autofoszforilációnak nevezik.
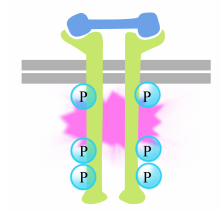
A mőszer a tyrosines a receptor írás kiváltja a közgyűlés egy intracelluláris jelátviteli komplex az írás. Az újonnan foszforilált tirozinok kötőhelyként szolgálnak a fehérjék jelzésére, amelyek ezután továbbadják az üzenetet más fehérjéknek. Egy fontos fehérjét, amelyet később a tirozin-kináz receptor jelző komplexei aktiválnak, Ras-nak nevezzük.
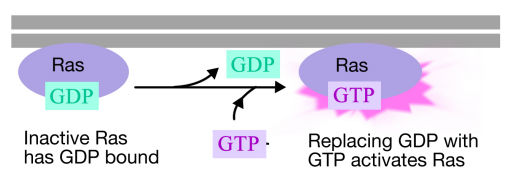
a RAS fehérje egy monomer guanin nukleotidkötő fehérje, amely a plazmamembrán citoszolos felületéhez kapcsolódik (valójában nagyon hasonlít a trimerikus g-fehérjék alfa alegységéhez). Csakúgy, mint egy g – fehérje alfa alegysége, a RAS akkor aktív, amikor a GTP kötődik hozzá, és inaktív, amikor a GDP kötődik hozzá.Továbbá, mint az alfa alegység, a RAS hidrolizálja a GTP-t a GDP-hez.
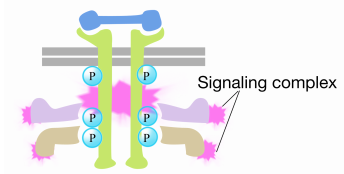
amikor egy jel érkezik a receptor tirozin-kinázhoz, a receptor monomerek összegyűlnek, és egymás tirozinjait foszforilálják, kiváltva a fehérjék komplexének összeszerelését a receptor citoplazmatikus farkán. Az egyik fehérje ebben a komplexben kölcsönhatásba lép a Ras-szal, és serkenti az inaktív Ras-hoz kötött GDP cseréjét egy GTP-hez. Ez aktiválja a Ras-t.
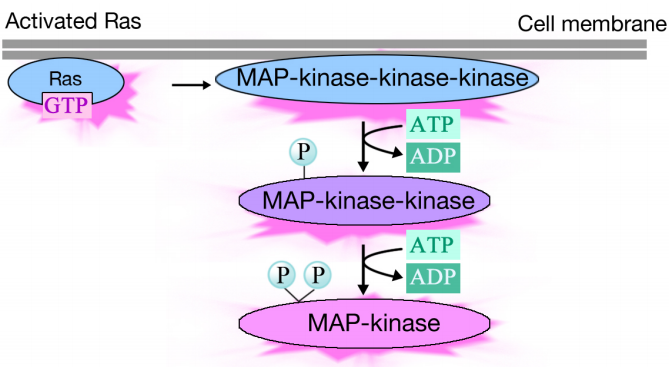
Az aktivált Ras három protein kináz foszforilációs kaszkádját váltja ki, amelyek továbbítják és terjesztik a jelet. Ezek a protein kinázok a MAP kinázok (mitogén aktivált Protein kinázok) nevű csoport tagjai. A végső kináz ebben a kaszkádban foszforilálja a különböző célfehérjéket, beleértve az enzimeket és transzkripciós aktivátorokat, amelyek szabályozzák a gén expresszióját.
a különböző enzimek foszforilációja megváltoztathatja tevékenységüket, és új kémiai reakciókat indíthat el a sejtben, míg a transzkripciós aktivátorok foszforilációja megváltoztathatja, hogy mely gének expresszálódnak. A génexpresszió és a fehérje aktivitás változásainak együttes hatása megváltoztatja a sejt élettani állapotát.
ismét, az RTKs által közvetített jelátvitel útját követve, meg lehet különböztetni az események ugyanazon alapvető mintáját: a jelet a receptor tirozin-kinázok extracelluláris tartományai kötik össze, ami a receptor dimerizációját és a citoszolos farok autofoszforilációját eredményezi, így közvetíti az üzenetet a sejt belsejébe.
az üzenetet jelzőkomplexumon keresztül továbbítják a RAS-nak, amely ezután egy sor kinázt stimulál. A kaszkád terminális kináza a célfehérjékre hat, és változásokat idéz elő a fehérje aktivitásában és a gén expressziójában.
A fenti leírások nyújt egy nagyon egyszerű vázlatot néhány főbb osztályok receptorok foglalkozni elsősorban a mechanisztikus részleteket a lépéseket, amelyek révén kapott jelek a különböző típusú receptorok változásokat a sejtek. A fő take-home lecke a különböző utak alapvető hasonlósága.
egy másik szempont, amelyet szem előtt kell tartani, az, hogy miközben minden egyes útvonalat elszigetelten néztünk, egy sejt bármikor több jelet kap, amelyek egyszerre különböző válaszokat indítanak el. A fent leírt útvonalak jelentős mértékű “keresztbeszélgetést” mutatnak, és az adott jelre adott választ a sejt által egyidejűleg fogadott egyéb jelek befolyásolják. A különböző receptorok, jelek sokasága és ezek kombinációi azok az eszközök, amelyekkel a sejtek képesek reagálni a különböző körülmények hatalmas változatosságára.
-
Dr. Kevin Ahern és Dr. Indira Rajagopal (Oregoni Állami Egyetem)
Leave a Reply