mi befolyásolja a csoportok méretét, amelyekben a főemlősök úgy döntenek, hogy élnek?

a főemlősök tanulmányozásának egyik legfontosabb elméleti hozzájárulása annak megértése, hogy mi befolyásolja a társadalmi struktúra és szervezetek komplex változásának természetét. Ennek hátterében egy sor elméleti fejlemény áll, amelyek azt vizsgálják, hogy az állatok miért élnek elsősorban csoportokban, és miért vannak bizonyos méretű csoportok. Itt arra a kérdésre összpontosítunk ,hogy ” mi befolyásolja a főemlősök által választott csoportok méretét?”
a főemlősök különösen hasznosak a kérdés megválaszolásához, mivel csoportméretük rendkívül változó, nem csak a fajokon belül, hanem a fajok között is. A vörös colobus (Procolobus rufomitratus) fajspecifikus csoportmérete például 12-150 tag között változik (Chapman & Chapman 2000a). Az interspecifikus variáció egyetlen egyéntől származik, amint azt az orangutánok (Pongo sp.) (van Schaik 1999), több mint 800 tagja mandrills (Mandrillus sphinx) (Abernethy et al. 2002). Ezenkívül egyes fajokon belül a társadalmi csoportok az idő múlásával többször is különböző méretű és kombinációs alcsoportokba osztódnak és újra egyesülnek (például a pókmajmok hasadási-fúziós társadalmi szervezete (ateles sp.) és a csimpánzok (Pan troglodytes), illetve a gelada (Theropithecus gelada) és a Hamadryas páviánok (Papio hamadryas, Aureli et al. 2008). Ez a variáció alapot nyújt a kutatók számára olyan modellek kidolgozásához, amelyek mind a csoportméret ökológiai, mind társadalmi mozgatórugóit vizsgálják.
a Csoportosítás több szempontból is előnyös. Egyének nagyobb csoportok hittem, hogy csökkent a kockázata a ragadozók (Hamilton 1971), lehet, hogy jobb találni, hogy megvédje az élelmiszer-forrásokat (Cody 1971, Wrangham 1980), de lehet ellen védett conspecific fenyegetés, mint a gyermekgyilkosság által extra-csoport, férfiak (Wrangham 1979). Különböző kutatók azt javasolták, hogy a csoportosítás ilyen kiszámítható előnyöket biztosít (Alexander 1974, van Schaik 1983), hogy a csoportméret különbségei a hátrányokkal magyarázhatók (Wrangham et al. 1993). A csoportosítás legszélesebb körben elfogadott lehetséges költsége a takarmányozási hatékonyság csökkenése. Hogy más egyének az azonos táplálkozási követelményeknek, az azt jelenti, hogy az állatok sem veszekedni élelmiszer (verseny, verseny), vagy egy állat, egy csoport veri a másik, hogy az étel, ezért ha a második állat jön egy olyan területre, egyszerűen nincs élelmiszer-bal (habar verseny, Janson & van Schaik 1988). Mindkét esetben úgy gondolják, hogy az élelmiszerekkel szembeni verseny azt eredményezi, hogy az állatoknak tovább kell utazniuk. Ennek az érvnek a logikája viszonylag egyszerű. Az állatoknak olyan területen kell takarmányozniuk, amely megfelel az energetikai és táplálkozási igényeiknek. Ebből következik, hogy a csoportméret növekedése növeli azt a területet, amelyet fedezni kell a megfelelő élelmiszer-ellátás megtalálásához. Így az egyéneknek tovább kell utazniuk, és több energiát kell fordítaniuk, ha nagy csoportban vannak, mint ha kisebb csoportban táplálkoznak. Az utazásra fordított idő növekedésével egy olyan pont kerül megközelítésre, ahol az utazásra fordított energia túl költséges, és a kisebb csoportok előnyössé válnak. Ily módon az ökológiai tényezők befolyásolhatják a mozgási mintákat és a takarmányozási hatékonyságot, ezáltal korlátozva a rendelkezésre álló élelmiszer-erőforrásokat hatékonyan kihasználó csoportok méretét. Ezeket az elképzeléseket az úgynevezett ökológiai korlátok modelljében formalizálták (Chapman & Chapman 2000B, Ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
az ökológiai korlátok modelljének lényeges eleme, hogy a csoportméret növelésének a csoporton belüli etetési verseny növekedéséhez kell vezetnie. Elképzelhető, hogy ez kissé eltérő módon működik, az egyes fajok által használt erőforrások jellegétől függően. A frugivorous, és esetleg sok folivorous főemlősök, hogy a takarmány diszkrét foltok-jellemzően fák hordozó élelmiszer tételek vagy csomókat a fák – További csoport tagjai kimerítik foltok gyorsabb és vezet fokozott nap tartományok (Chapman 1988, Snaith & Chapman 2005). Több rovar faj, amelynek forrásai nem fordulhat elő, mint különálló foltok, folyamatos utazás során a lombkorona keresés a rovarok további csoport tagjai növekedéséhez vezethet az átfedés az egyes keresési mezők, csökkenti az egy főre jutó találkozás árak ételekkel vagy így növelve a területen kell keresni (van Schaik et al. 1983).
tehát az ökológiai korlátok modellje azt sugallja, hogy azok a tényezők, amelyek befolyásolják az állatok utazásának távolságát, szintén befolyásolják a csoport méretét. Azoknál az állatoknál, amelyek jellemzően gyümölcsökkel vagy levelekkel táplálkoznak, amelyek kimerülhetnek, a tapasz mérete határozza meg, hogy egy adott méretű csoport mennyi ideig maradhat és táplálkozhat. Egy nagy csoport kevesebb időt töltene egy adott méretű javításban, mint egy kisebb csoport, mert gyorsabban kimeríti a tapaszt — egy nagy csoportnak egyszerűen több szája van a takarmányozáshoz. Ha az állatok a tapaszok között utaznak, miután kimerítették őket, akkor a tapaszok sűrűsége és eloszlása határozza meg a felmerült utazási költségeket. Ha az erőforrás-javítások nagy sűrűségűek vagy összetapadt eloszlásban vannak, a következő tapasztól való távolság kicsi, az utazási költségek alacsonyak, ezért az állatok nagy csoportokat képezhetnek. Ilyen esetekben a nagy csoport tagságával járó többletköltségek, például a sok javítás meglátogatásának szükségessége könnyen behajthatók. Ezzel szemben, amikor az erőforrás-javítások alacsony sűrűségnél fordulnak elő, a következő tapasztól való távolság általában nagy, az utazási költségek magasak, az állatok nem engedhetik meg maguknak a foltok gyors kimerülését, ezért az állatok kis csoportokat alkotnak.
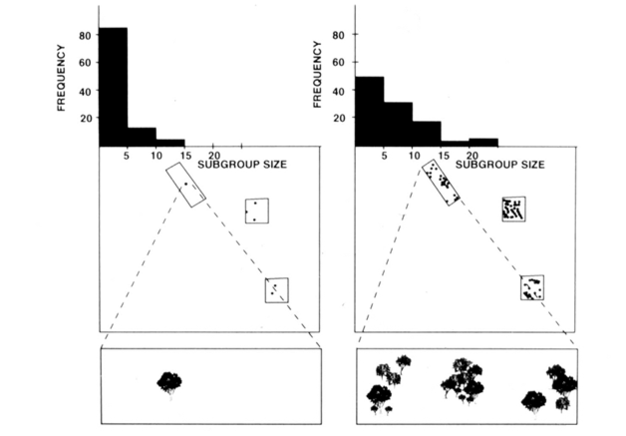
A fissziós-fúziós társadalmi szervezet a pókmajom, csimpánzok, valamint néhány más főemlősök (1. Ábra) kínál hasznos vizsgálatok az ökológiai korlátok modell, mert megpróbálja megjósolni, időbeli változások alcsoport méret a közvetlen intézkedések a mérete, sűrűsége, valamint az élelem elosztása a források (Chapman 1990, Chapman et al. 1995). A pókmajmok és a csimpánzok esetében egy többszörös regressziós modellt fejlesztettek ki annak bizonyítására, hogy a havonta mért élelmiszer-tapaszok mérete, sűrűsége és eloszlása jó előrejelzője volt az alcsoport méretének. Nehéz vizuálisan ábrázolni a többszörös regressziót, de a pókmajmokhoz használt módszereket a 2.ábrán ábrázolják. A pókmajmok nagyon rugalmas hasadási fúziós típusú társadalmi szervezettel rendelkeznek, a Costa Rica-i Santa Rosa Nemzeti Parkban pedig az alcsoport mérete 1-35 személy lehet, de átlagosan körülbelül 5 egyedet találnak együtt utazva. Ebben a populációban az átlagos havi alcsoportméret varianciájának 50% – a megjósolható az élelmiszerfoltok méretének, sűrűségének és eloszlásának viszonylag durva méréseiből (Chapman 1990).

a fent említett fajok mellett az ökológiai korlátok modell alkalmazhatóságát általában különféle fajokból származó kutatások támogatták (Snaith & Chapman 2005) és helyzetek (pl. általános modellek, Wrangham et al. 1993 és vegyes fajtársaságok, Chapman & Chapman 2000C). Úgy véljük, hogy tekintettel arra a széles körű támogatásra, amelyet ez a modell kapott, eljött az idő, hogy ésszerű azt sugallni, hogy a fajoknak általában meg kell felelniük az ökológiai korlátok modell elvárásainak. Ez nem jelenti azt, hogy minden faj meg fog felelni, így izgalmas időszak lesz a kutatás ezen a területen, mert a kutatók kivételeket kereshetnek a modelltől. Gyanítjuk, hogy a nem megfelelő fajok ezt megteszik, mert olyan társadalmi stratégiákat fogadtak el, amelyek ellentétesek az ökológiai elvárásokkal. Ez “mércét” jelent a különböző társadalmi stratégiák potenciális jelentőségének értékeléséhez (azaz a további eltér az ökológiai modell elvárásaitól, annál fontosabb a társadalmi szelektív nyomás).
röviden mutassunk be két példát korábbi kutatásainkból. A nőstény csimpánzok magányosabbak voltak, mint a nőstény pókmajmok, mégis mindkettőjüknek hasonló hasadási fúziós társadalmi szervezetei vannak. Továbbá, miközben pontosan meg tudtuk jósolni a hímek és aladult csimpánzok számát egy alcsoportban az ökológiai feltételek alapján, nem tudtuk megjósolni a női csimpánzok számát ezekben az alcsoportokban (Chapman et al. 1995). Még akkor is, ha az erőforrások rendkívül bőségesek voltak, és szinte az összes hím egy nagy alcsoportban volt, a nőstények ritkán léptek be csoportokba, ami arra utal, hogy egy nagyobb alcsoportban való tartózkodás költsége meghaladja az előnyöket, mint például a ragadozók elkerülése. Ez váratlan abból a szempontból, hogy ezeknek a nőstényeknek a csecsemői az életkor/nemi osztály lenne, amelyet leginkább az predáció fenyeget. Az egyik tesztelhető hipotézis ezeknek a megfigyeléseknek a magyarázata az, hogy a két fajban a koalíciók jellege befolyásolhatja a csoporttagság előnyeit. A bizonyítékok arra utalnak, hogy a csimpánzokkal ellentétben a pókmajom nőstények olyan koalíciókat alkotnak, amelyek gyakran működnek, hogy a koalíció tagjai kizárólagos hozzáférést biztosítsanak az élelmiszerekhez. Tehát a pókmajmok, amikor csatlakoznak bizonyos egyénekhez, növelhetik az élelmiszerhez való hozzáférést mások kizárásával, de a csimpánz nőstények esetében ez nem így van (Chapman et al. 1995).
azonosítottunk egy másik érdekes eltérés, potenciálisan által okozott társadalmi befolyásoló tényezők, a költségek, valamint előnye, hogy egy csoport, s ez vonatkozik kolobusz, fekete-fehér kolobusz majmok (aka. guerezas, Colobus guereza) Az Ugandai Kibale Nemzeti Parkban. Vörös colobus alkotnak nagy csoportok átlagosan 65 egyének (25-127 egyének, Snaith et al. 2008), míg a guerezas kis csoportokban él, átlagosan 6, 5 egyének (4-11 egyének, Harris & Chapman 2007). E különbség ellenére a két faj által fogyasztott növényekben nagy a hasonlóság. Ha az ökológiai feltételek felelősek lennének a két kolobinfaj közötti csoportméret különbségéért, akkor azt várnánk, hogy étrendjük eltérne, és az élelmiszerfák sűrűsége alacsonyabb lenne a guerezák otthoni tartományaiban, mivel kisebb csoportméretük van. Étrendjük azonban nagyon hasonló; a Red colobus és a guereza csoport étrendi átfedése, amely teljes egészében a red colobus csoport otthoni tartományában volt, 43 volt.2%, míg a két szomszédos csoport vörös colobus, diéta átfedésben csak 37.3% (Chapman et al. 2002). Úgy tűnik, hogy a női guereza reproduktív siker maximalizálódik a kis és közepes méretű csoportok, vagy azért, mert a nagyobb csoportok tapasztalnak magasabb aránya átvételi és infanticid vagy több etetés verseny (lásd bizonyíték tülekedés verseny a folivorous Trachypithecus phayrei és Semnopithecus sp. , de a bizonyítékok hiánya guerezas kimerítő élelmiszer foltok ). Számos férfi átvétel és csecsemőgyilkosság szemtanúi voltunk guerezasban (Onderdonk 2000) – lásd Teichroeb & Sicotte (2008) az infanticidre vonatkozó elmélet értékeléséhez a kolobinokban. Nem világos, hogy az infanticid kockázata hogyan korlátozza a guereza csoportok méretét, amikor nyilvánvaló hozzáféréssel rendelkeznek egy nagy sűrűségű élelmiszerhez, amely lehetővé teszi a több hímes csoportok kialakulását. Több fajnál a nagyobb nősténycsoportok inkább a vándorló hímek célpontjai, és ez a helyzet növelheti az infanticidek arányát-például a Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & Van Schaik 2001). Azonban gyermekgyilkosság várhatóan kevésbé gyakori, ha egy csoport tartalmaz több szaporodást aktív férfiak, mert a férfi bevándorlási, valamint az azt követő erőszakos pedig gyermekgyilkosság kevésbé fordulhat elő, ha a jövevény szembe több lehetséges úrrá a jövő csecsemők (Janson & van Schaik 2000). Guerezas úgy tűnik, hogy bőséges élelmiszer-források alkotnak nagyobb csoportok (Tombak et al. 2012), ami csökkentené az infanticid kockázatát, mégis nem alkotnak nagy csoportokat. Lehetséges, hogy a férfi-férfi kölcsönhatások jellege guerezasban a nagy csoportokat instabillá teszi. Úgy tűnik, ez a helyzet medveféle kolobusz (Kolobusz vellerosus), ahol a multi-férfi csoportok vonzza a férfi bevándorlók, illetve magasabb gyermekgyilkosság árak, mint a csoportok egy erős felnőtt férfi, miniszterelnök (Teichroeb, Wikberg, Badescu, & Sicotte, nem publikált adatok; 3.Ábra). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

Ha az ökológiai korlátok modell felvázolja nyomás, hogy általában befolyásolja a prímás csoport mérete, illetve, ha az ettől való eltéréseket várható csoport mérete eredménye, társadalmi nyomás, lesz egy izgalmas időszak ez a kutatási terület, mert felvet néhány érdekes kérdést. Például: Milyen jellegű a társadalmi nyomás a csoportméretre? Miért vannak jelen ezek a társadalmi korlátok ezekben a fajokban, és miért nem kapcsolódnak szorosan egymáshoz? És mit választanak az evolúciós erők ezekre a társadalmi nyomásokra? A csoportméret kritikus elem, amely befolyásolja az elfogadott társadalmi szervezet típusát, így ezeknek a kérdéseknek a vizsgálata a szocioekológiai modellek izgalmas finomításához vezet.
Leave a Reply