Morula Fejlesztési
a Google Translate – válassza ki a nyelvet a listából az alább látható (ez megnyit egy új külső oldal)
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | fülöp-szigeteki | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский / Español / szuahéli / Svensk / ไทย / Türkçe / اردو /יייי / Tingng Việt ezek a külső fordítások automatizáltak és nem pontosak. (Még? Arról, Fordítások)
Bevezető
(Latin, morula = mulberry) korai szakaszban a poszt-trágyázás fejlesztési amikor a sejtek gyorsan mitotically osztva, hogy készítsen egy szilárd tömege sejtek (12-15 sejtek) a “mulberry” megjelenése. Ezt a stádiumot egy üreg kialakulása követi ebben a celluláris tömeg blasztocisztás szakaszban.
A legfontosabb esemény előtt morula formáció “tömörítés”, ahol a 8 sejtes embrió változáson megy keresztül a sejtek morfológiája, illetve sejt-sejt adhézió, hogy kezdeményezi a kialakulását ez a szilárd labdát a sejtek.
emberben a morula fejlődési stádiuma a megtermékenyítést követő első hét első napjaiban (GA 3.hét) fordul elő, és a Carnegie 2. fázisa. Ezt a lépést egy üreg, a blastocoel kialakulása követi, amely meghatározza a blastociszta kialakulását.

ART Preimplantációs blasztomer biopszia
|
Az Asszisztált Reprodukciós Technológia, a morula stádiumban, amikor az egyik legkorábbi prenatális diagnosztikai vizsgálatot lehet végezni, azáltal, hogy megszünteti egy cella (blasztomer), valamint végző genetikai diagnózis a DNS-t. |
Molekuláris – az egér, átmenet során a morula, hogy blastocyst színpadon, a differenciálás, a belső cella-tömeg (ICM), valamint trophectoderm (TE) kimutatták, hogy szabályozza a Víziló út.
Morula linkek: morula / Carnegie 2. szakasz / mitosis / blastocist / megtermékenyítés / 1. hét / előadás 1. hét / Kategória: Carnegie 2. szakasz | Kategória:Morula
Néhány újabb Megállapítások
- Hatása Downregulating a Víziló Út Tagok YAP1, valamint LATS2 Átiratok a Korai Fejlesztés, valamint a génexpresszió vett Részt a Differenciálás a Sertés Embriók “egér fejlődés, a differenciálás, a belső cella-tömeg (ICM), valamint trophectoderm (TE) az átmenet során a morula, hogy blastocyst szakaszban szabályozza a Víziló út; azonban a funkciók a Víziló út a sertés embryogenesis nem vizsgálták. A jelen tanulmányban megvizsgáltuk a génexpresszió minták a Víziló út tagok igen-asszociált fehérje 1 (YAP1), illetve nagy tumor szupresszor 2 (LATS2), valamint a funkciók ezek a gének során sertés preimplantációs fejlesztési segítségével RNS interferencia. Mind a YAP1, mind a LATS2 mRNS-szint magas volt az in vitro Érett petesejtekben és az 1-sejtes stádiumú embriókban, és fokozatosan csökkent a fejlődéssel. A yap1 nukleáris lokalizációját a morula és a blasztociszta szakaszaiban észlelték. A yap1 vagy a LATS2 csökkenése gátolta a sertés preimplantációs fejlődését, és befolyásolta a Pou 5.osztályú homeobox 1 (OCT-4) és a SRY-vel kapcsolatos HMG-box gén 2 (SOX2) expressziós szintjét, az ICM/TE differenciáláshoz szükséges transzkripciós tényezőket. A yap1 és a LATS2 együttesen elengedhetetlen a sertéspreimplantáció fejlesztéséhez, és lehetséges, hogy a Vízilóút fontos szerepet játszik a sertés ICM/TE szegregációban.”
- az emberi és egér homeobox gén expressziójának sokfélesége a fejlődésben és a felnőtt szövetekben ” 8-sejt-morula, 12 eutherian-specifikus homeobox gén, amely nem mutatható ki a reproduktív szöveteken vagy az embrión kívül (ROXF2, ROXF2B, CPHX1, CPHX2, DPRX, LEUTX, TPRX1, TPRX2, ARGFX, NANOGNB, DUXA, DUXB). Hox

Ez a táblázat lehetővé teszi a külső PubMed adatbázis automatizált számítógépes keresését a felsorolt “keresési kifejezés” szöveges link segítségével.
- ez a Keresés most kézi linket igényel, mivel az eredeti PubMed kiterjesztés le van tiltva.
- a megjelenített hivatkozások listája nem tükrözi a tartalom vagy relevancia alapján az anyagok szerkesztői kiválasztását.
- referenciák is megjelennek ezen a listán a tényleges oldalnézet dátuma alapján.
A tartalom oldal többi részén és a kapcsolódó vitafórumon (a kiadvány év alcímei alatt felsorolva) felsorolt hivatkozások tartalmaznak néhány szerkesztői választást, mind a relevancia, mind a rendelkezésre állás alapján.
Több? Referenciák | Vita Oldal | Lap Keresések | 2019 Referenciák | 2020 Hivatkozások
Keresés kifejezés: Morula Fejlesztés | Morula Tömörítés | Morula Blasztomer Osztály | Morula Víziló |
Ezek a papírok eredetileg megjelent a Néhány újabb Megállapítások asztalra, de mint azt a listát nőtt a hossza már csoszogott le, hogy ez az összecsukható asztal.
Lásd még a vitafórumot az évenként felsorolt egyéb hivatkozásokról, valamint az aktuális oldalon található hivatkozásokról.
- Par-aPKC-függő, illetve független mechanizmusok együttműködve kontroll sejt polaritás, Víziló jelzés, illetve sejt elhelyezése a 16-mobil színpad egér embriók “A preimplantációs egér embriók, a Víziló jelátvitelben központi szerepet játszik a szabályozó a sorsa a trophectoderm (TE), valamint a belső cella-tömeg (ICM). A több mint 32 sejtes korai blasztocisztákban a par-aPKC rendszer szabályozza a külső sejtek polarizációját az apicobasalis tengely mentén, a sejtpolaritás pedig elnyomja a Hippo jelátvitelt.”Víziló | egér
- a Nem-invazív képalkotó az emberi embriók előtt embrionális genom aktiválás jósolja, fejlesztési, hogy a blastocyst színpadon, hogy “Mi a jelentés tanulmányok a preimplantációs emberi embrió fejlődése, amely korrelál a time-lapse képek elemzése, valamint a génexpresszió profilalkotás. Az in vitro megtermékenyítésből (IVF) származó nagy zigóták vizsgálatával megállapíthatjuk, hogy a blastociszta szakaszba való progresszió sikere a >93% érzékenységgel és specificitással megjósolható, három dinamikus, nem invazív képalkotó paraméter mérésével a megtermékenyítés utáni 2.napra, az embrionális Genom aktiválása (EGA) előtt.”
- a Funkcionális genomika 5 – 8-mobil színpad emberi embriók által blasztomer egysejtű cdns elemzés “negyvenkilenc blastomeres 5-től – 8-sejtes embrió vizsgálták következő hatékony egysejtű cdns hangerősítő jegyzőkönyv biztosítani a sablon, nagy sűrűségű microarray analízis. A korábban leírt markerek, jellemző a Belső Cella-Tömeg (ICM) (n = 120), stemness (n = 190), valamint Trophectoderm (TE) (n = 45), elemeztük, valamint egy háztartási minta 46 gének állapították meg. …Összefoglalva, az 5-8 sejtes emberi embriók globális egysejtes cDNA amplifikációs mikroarray elemzése azt mutatja, hogy a blastomere sors nem elkötelezett az ICM vagy a TE iránt.”
Movies
|
Morula Model
p > page | Play
|
Zygote Mitosis Page | Play
|
Early Division Page | Play
|
Parental Genomes Page | Play
|
Mouse Blastocyst Page | Play
|
Movies
Compaction
- E-cadherin mediated adhesion initiates at compaction at the 8-cell stage
- regulated post-translationally via protein kinase C and other signalling molecules
Blastomere Division

a humán blasztociszták fejlődésének in vitro vizsgálata azt mutatta, hogy azok a blasztomerek, amelyek kezdetben gyorsan osztódnak, nagyobb valószínűséggel alakulnak ki blasztociszta szakaszba.
egy nemrégiben egereken végzett vizsgálat azt mutatta, hogy a mitotikus orsó nem volt specifikus orientáció a sejtosztódás során a 8-16 sejtszakasz átmenet során. Ez arra utal, hogy a 8 sejtes szakaszban nincs előre meghatározott hasítási mintázat (előre mintázott), amelyet csak a mitózis során a sejtek kerekítésének mértéke modulál. Más fajokban, például a C féregben.az elegans és az ascidians a zigóta szakaszból sajátos orsó-orientációs mintázattal rendelkeznek.
Model Human Morula Development
az alábbi ábra egy nemrégiben készült, a megtermékenyítést követő 1. héten az in vitro humán fejlődés video-és genetikai elemzését alkalmazó vizsgálatból származik.
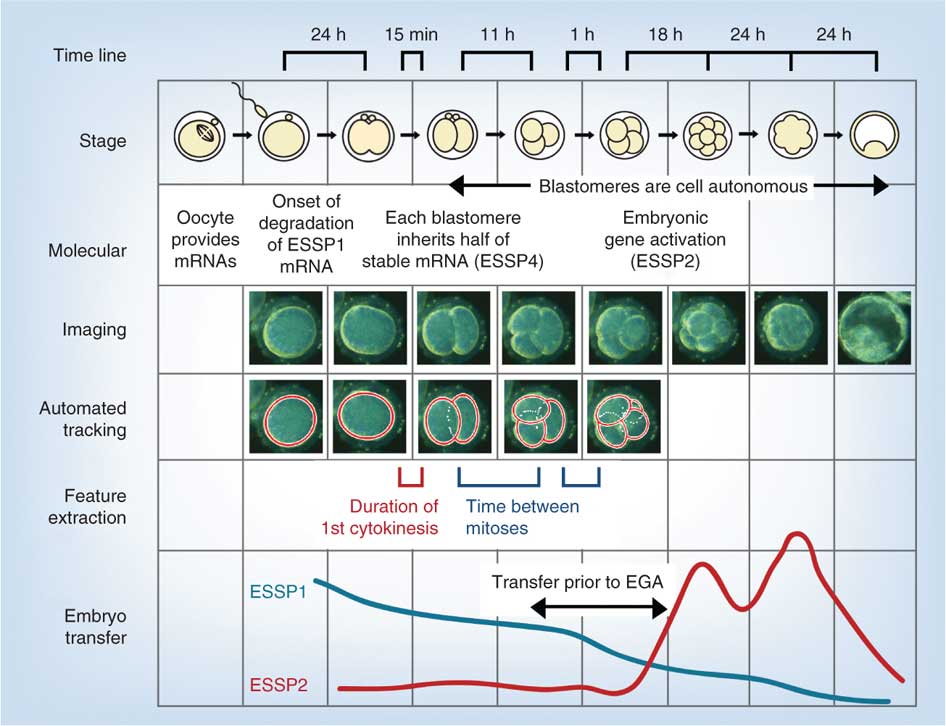
- EGA-embrionális Genom aktiválás
- ESSP-embrionális stádiumspecifikus minta, négy egyedi embrionális stádiumspecifikus mintázat (1-4)
linkek: A
más fajokban előforduló Morulák
egér Morula
az egérben a moruláról blastocisztára való átmenet során kimutatták, hogy a belső sejttömeg (ICM) és a trophectoderm (TE) differenciálódását a Hippo útvonal szabályozza.
-

4 cell morula stage development
-

Sox2 expression
-

Early gene expression
-

Early gene expression
-

Early gene expression
-

Early génexpresszió
linkek: Egérfejlődés
Tengeri Sün Morula

Tengeri Sün korai embrió hasítási mintája (SDB Galéria Képek)
linkek: Tengeri Sün fejlesztés
szarvasmarha Morula
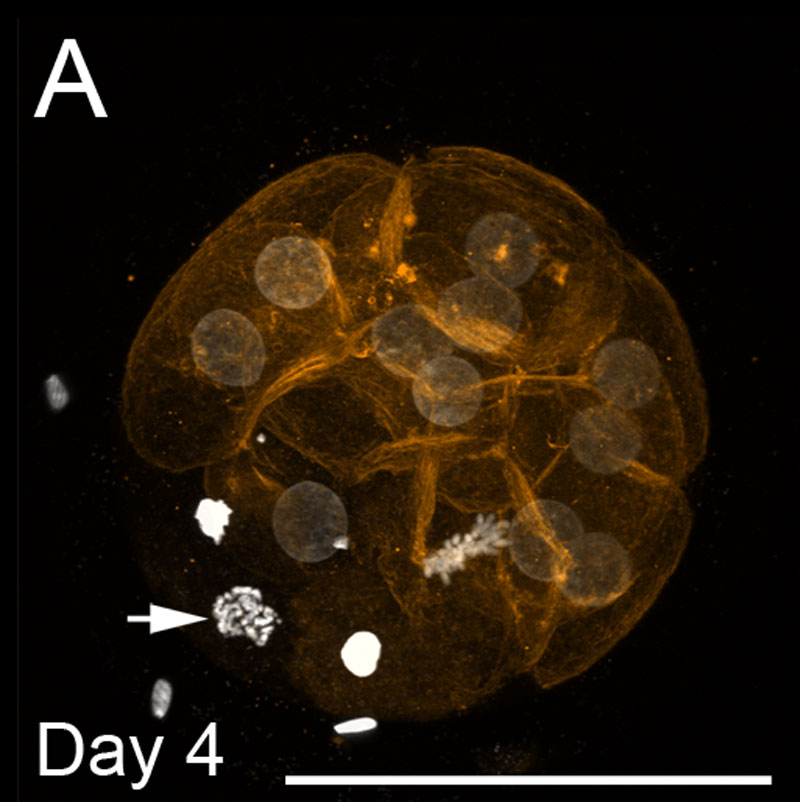
szarvasmarha Morula
- a képen a DNS-festés (fehér) és az F-aktin szálak (narancssárga) látható a 4.napon. A méretarányok 100 µm-t képviselnek.
- a halvány színű kerek magok interfázisban vannak.
- a nyíl egyetlen magot mutat a prophase-ban.
- a metafázisban egyetlen mag látható.
- a kondenzált fényes magok apoptotikusak.
linkek: szarvasmarha fejlődés / mitózis
Morula biopszia

kompakt morula-stádiumú embriók biopsziája
- (a) egy kompakt morula-stádiumú embrió biopszia előtt.
- (B–G) a biopszia lépései.
- (H) embrió 2 órával a biopszia után.
linkek: prenatális diagnózis / asszisztált reprodukciós technológia
- 1, 0 1.1 Zhang P, Zucchelli M, Bruce S, Hambiliki F, Stavreus-Evers A, Levkov L, Skottman H, Kerkelä E, Kere J & Hovatta O. (2009). Az emberi implantáció előtti fejlődés transzkriptóm profilozása. PLoS ONE, 4, e7844. PMID: 19924284 DOI.
- Milacsics T. (2014). A preimplantáció és a prenatális genetikai szűrés, valamint a nem invazív vizsgálatok új eredményei, mint a csecsemők egészségi állapotának potenciális előrejelzője. Biomed Res Int, 2014, 306505. PMID: 24783200 DOI.
- 3.0 3.1 Hirate Y, Hirahara S, Inoue K, Kiyonari H, Niwa H & Sasaki H. (2015). A Par-aPKC-függő és-független mechanizmusok együttműködően szabályozzák a sejtek polaritását, a Hippo jelátvitelt és a sejtek pozicionálását a 16 sejtes stádiumú egér embriókban. Dev. A Növekedés Különbözik. , 57, 544-56. PMID: 26450797 DOI.
- 4.0 4.1 4.2 Emura N, Saito Y, Miura R & Sawai K. (2020). A Hippo Pathway tagjai Yap1 és LATS2 transzkripciójának a korai fejlődésre és gén Expresszióra gyakorolt hatása, amely részt vesz a Sertésembriók Differenciálásában. A Sejt Újraprogramozása,,. PMID: 32150685 DOI.
- Dunwell TL & Holland PW. (2016). Az emberi és egér homeobox gén expressziójának sokfélesége a fejlődésben és a felnőtt szövetekben. BMC Dev. Biol. , 16, 40. PMID: 27809766 DOI.
- 6.0 6.1 Wong CC, Loewke KE, Bossert NL, Behr B, De Jonge CJ, Baer TM & Reijo Pera RA. (2010). Az emberi embriók nem invazív képalkotása az embrionális Genom aktiválása előtt előrejelzi a fejlődést a blasztociszta szakaszába. Nat. Biotechnol. , 28, 1115-21. PMID: 20890283 DOI.
- Galán A, Montaner D, Póo ME, Valbuena D, Ruiz V, Aguilar C, Dopazo J & Simón C. (2010). Az 5-8 sejtes emberi embriók funkcionális genomikája blastomere egysejtű cDNA analízissel. PLoS ONE, 5, e13615. PMID: 21049019 DOI.
- 8, 0 8, 1 Dard N, Louvet-Vallée S & Maro B. (2009). A mitotikus orsók orientációja az egér embriókban a 8-16 sejtes átmenet során. PLoS ONE, 4, e8171. PMID: 19997595 DOI.
- Fenwick J, Platteau P, Murdoch AP & Herbert M. (2002). A megtermékenyítéstől az első hasításig eltelt idő az emberi preimplantációs embriók fejlődési kompetenciáját jósolja in vitro. Zümmög. Feddés. , 17, 407-12. PMID: 11821286
- Leidenfrost S, Boelhauve M, Reichenbach M, Güngör T, Reichenbach HD, Sinowatz F, Wolf e & Habermann FA. (2011). Sejtmegállás és sejthalál az emlős előimplantáció fejlődésében: a szarvasmarha-modell tanulságai. PLoS ONE, 6, e22121. PMID: 21811561 DOI.
- Zakharova EE, Zaletova VV & Krivokharchenko AS. (2014). Az emberi morula-stádiumú embriók biopsziája: 215 IVF/ICSI ciklus eredménye PGS-vel. PLoS ONE, 9, e106433. PMID: 25191937 DOI.
Coticchio G, Lagalla C, Sturmey R, Pennetta F & Borini A. (2019). A rejtélyes morula: a fejlődés mechanizmusai, a sejt sorsának meghatározása, az önkorrekció és a művészetre gyakorolt hatások. Zümmög. Feddés. Frissítés: 25, 422-438. PMID: 30855681 DOI.
Bessonnard S, Mesnard D & Constam DB. (2015). A PC7 és a kapcsolódó proteázok, a Furin és a Pace4 szabályozzák az e-cadherin funkciót a blasztociszták kialakulása során. J. Cell Biol. , 210, 1185-97. PMID: 26416966 DOI.
Dzamba BJ, Jakab KR, Marsden M, Schwartz MA & DeSimone DW. (2009). Cadherin adhézió, szöveti feszültség, nem kanonikus Wnt jelátvitel szabályozza fibronektin mátrix szervezet. Dev. Cella , 16, 421-32. PMID: 19289087 DOI.
Santos J, Pereira CF, Di-Gregorio A, lucfenyő t, éger O, Rodriguez T, Azuara V, Merkenschlager M & Fisher AG. (2010). A pluripotens és az embrionális őssejtek epigenetikai és újraprogramozási tulajdonságaiban mutatkozó különbségek a kromatin átalakulását jelentik, ami fontos korai esemény a fejlődő egér embrióban. Epigenetika Kromatin, 3, 1. PMID: 20157423 DOI.
keresés PubMed
keresés Pubmed: morula development | blastomere development |
Szójegyzék linkek
Szójegyzék: a/B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | U | V | W | X | Y | Z | Numbers | term Link
idézzük ezt az oldalt: Hill, M. A. (2021, március 24) Embryology morula development. https://embryology.med.unsw.edu.au/embryology/index.php/Morula_Development
Leave a Reply