8.5: Receptor Tyrosine Kinases (RTKs)
Receptor tyrosine kinases mediano le risposte a un gran numero di segnali, inclusi ormoni peptidici come l’insulina e fattori di crescita come il fattore di crescita epidermico. Come i GPCR, le tirosin chinasi del recettore legano un segnale, quindi trasmettono il messaggio attraverso una serie di molecole intracellulari, l’ultima delle quali agisce sulle proteine bersaglio per modificare lo stato della cellula.
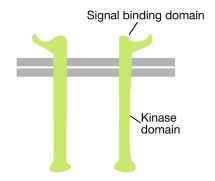
Come suggerisce il nome, un recettore tirosin chinasi è un recettore di superficie cellulare che ha anche un’attività tirosin chinasi. Il dominio di legame del segnale del recettore tirosin chinasi si trova sulla superficie cellulare, mentre l’attività enzimatica della tirosin chinasi risiede nella parte citoplasmatica della proteina (vedi figura sopra). Un’elica alfa transmembrana collega queste due regioni del recettore.
Cosa succede quando le molecole del segnale si legano alle tirosin chinasi del recettore?
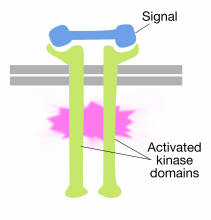
il legame di molecole segnale per i domini extracellulari dei recettori tirosina chinasi molecole provoca due molecole recettrici alla dimerizzazione (di riunirsi e di associarsi). Questo porta le code citoplasmatiche dei recettori vicini l’uno all’altro e fa sì che l’attività della tirosina chinasi di queste code sia attivata. Le code attivate si fosforilano a vicenda su diversi residui di tirosina. Questo è chiamato autofosforilazione.
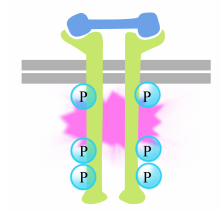
La fosforilazione delle tirosine sulle code dei recettori innesca l’assemblaggio di un complesso di segnalazione intracellulare sulle code. Le tirosine recentemente fosforilate servono come siti di legame per le proteine di segnalazione che poi passano il messaggio sopra ancora ad altre proteine. Una proteina importante che viene successivamente attivata dai complessi di segnalazione sulle tirosin chinasi del recettore è chiamata Ras.
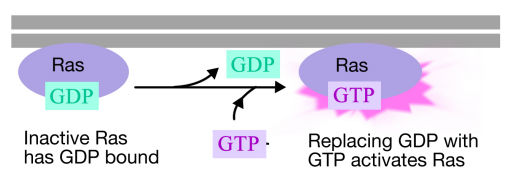
La proteina Ras è una proteina legante il nucleotide della guanina monomerica che è associata alla faccia citosolica della membrana plasmatica (infatti, è molto simile alla subunità alfa delle proteine G trimeriche). Proprio come la subunità alfa di una proteina G, Ras è attiva quando il GTP è legato ad esso e inattivo quando il PIL è legato ad esso.Inoltre, come la subunità alfa, Ras può idrolizzare il GTP in GDP.
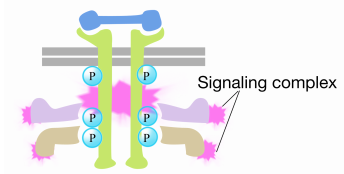
Quando un segnale arriva alla tirosina chinasi del recettore, i monomeri del recettore si uniscono e fosforilano le tirosine degli altri, innescando l’assemblaggio di un complesso di proteine sulla coda citoplasmatica del recettore. Una delle proteine in questo complesso interagisce con Ras e stimola lo scambio del PIL legato al Ras inattivo per un GTP. Questo attiva il Ras.
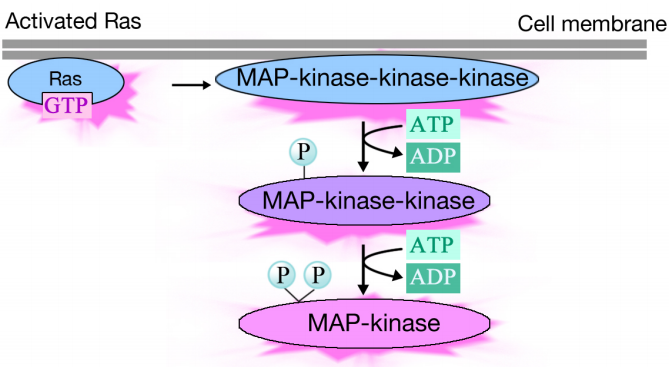
Attivato Ras innesca una cascata di fosforilazione di tre protein chinasi, che relè e distribuire il segnale. Queste protein chinasi sono membri di un gruppo chiamato MAP chinasi (Proteine chinasi attivate dal mitogeno). La chinasi finale in questa cascata fosforila varie proteine bersaglio, inclusi enzimi e attivatori trascrizionali che regolano l’espressione genica.
La fosforilazione di vari enzimi può alterare le loro attività e scatenare nuove reazioni chimiche nella cellula, mentre la fosforilazione degli attivatori trascrizionali può modificare i geni espressi. L’effetto combinato dei cambiamenti nell’espressione genica e nell’attività proteica altera lo stato fisiologico della cellula.
Ancora una volta, seguendo il percorso di trasduzione del segnale mediato da RTK, è possibile discernere lo stesso schema di base degli eventi: un segnale è legato dai domini extracellulari delle tirosin chinasi del recettore, con conseguente dimerizzazione del recettore e autofosforilazione delle code citosoliche, trasmettendo così il messaggio all’interno della cellula.
Il messaggio viene trasmesso tramite un complesso di segnalazione a Ras che stimola una serie di chinasi. La chinasi terminale nella cascata agisce sulle proteine bersaglio e provoca cambiamenti nelle attività proteiche e nell’espressione genica.
Le descrizioni di cui sopra forniscono uno schizzo molto semplice di alcune delle principali classi di recettori e trattare principalmente con i dettagli meccanicistici dei passaggi con cui i segnali ricevuti da vari tipi di recettori provocano cambiamenti nelle cellule. Una grande lezione da portare a casa è la somiglianza essenziale dei diversi percorsi.
Un altro punto da tenere a mente è che mentre abbiamo esaminato ogni singolo percorso in isolamento, una cellula, in un dato momento riceve più segnali che innescano una varietà di risposte diverse contemporaneamente. I percorsi sopra descritti mostrano un considerevole grado di” cross-talk ” e la risposta a un dato segnale è influenzata dagli altri segnali che la cellula riceve simultaneamente. La moltitudine di diversi recettori, segnali e le loro combinazioni sono i mezzi con cui le cellule sono in grado di rispondere a un’enorme varietà di circostanze diverse.
Collaboratori
-
Dr. Kevin Ahern e Dr. Indira Rajagopal (Oregon State University)
Leave a Reply