カマキリ
AnatomyEdit

カマキリは、くちばしのような鼻と下顎を持つ大きな三角形の頭を持っています。 彼らは2つの球根複眼、3つの小さな単純な目、そして一対の触角を持っています。 首の関節も非常に柔軟であり、いくつかの種のカマキリは頭をほぼ180°回転させることができます。 カマキリの胸郭は前胸郭、中胸郭、および中胸郭から構成されています。 Mantoida属以外のすべての種では、頭と前足を負う前胸は、他の2つの胸部セグメントよりもはるかに長い。 前胸はまた柔軟に連結され、ボディの残りが多かれ少なかれ不動に残る間、頭部および前部肢の動きの広い範囲を可能にする。 カマキリはまた、彼らは彼らの中胸部の聴覚室に二つの鼓膜で、鼓膜聴覚を持っているという点でDictyopteraに固有のものです。 ほとんどのカマキリは超音波のみを聞くことができます。
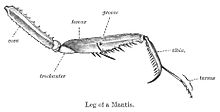
カマキリは、獲物のアイテムがキャッチされ、しっかりと保持されている二つのスパイク、把持前足(”raptorial脚”)を持っています。 カマキリの後四脚を含むほとんどの昆虫の脚では、coxaと転子は脚の目立たない基部として結合し、raptorial脚では、coxaと転子は、把持装置の棘のある部分である大腿骨と同じくらい長いセグメントを形成するために結合する(図を参照)。 大腿骨の基部には円盤状の棘があり、通常は4本の数であるが、種によってはどれも5本までの範囲である。 これらの棘の前には、脛骨に沿った同様の一連の結節およびその先端付近の頂端爪とともに、カマキリの前脚を獲物に与える多数の歯のような結節が 前足は歩行付属器として使用される繊細な足根で終わり、四つまたは五つのセグメントで構成され、アロリウムのない二本のつま先の爪で終わる。カマキリは、大翼(長い翼)、短翼(短い翼)、微小翼(痕跡翼)、またはapterous(翼のない)として緩く分類することができます。
カマキリは、巨大翼(長い翼)、brachypterous(短い翼)、微小翼(痕跡翼)、またはapterous(翼のない)。
カマキリは、 翼のない場合は、カマキリは翼の二組を持っています: 外側の翼、またはtegminaは、通常、狭くて革のようです。 それらはより明確、より敏感であるhindwingsのためのカムフラージュそして盾として作用する。すべてのカマキリの腹部は10のtergitesから成り、対応するセットは男性で目に見える9つのsternitesおよび女性で目に見える7つである。 腹部は女性よりも男性でスリムになる傾向がありますが、男女ともに一対のcerciで終わります。
VisionEdit

カマキリはステレオビジョンを持っています。 彼らは視力によって獲物を見つける;彼らの複眼は10,000ommatidiaまで含まれています。 中心窩と呼ばれる前部の小さな領域は、他の眼よりも視力が大きく、潜在的な獲物を調べるために必要な高解像度を生成することができます。 周辺ommatidiaは動きを感知することにかかわっています; 動いている物体に気付くと、頭部を急速に回転させて物体を中心窩の視野に持ち込む。 その後、獲物のさらなる動きは、カマキリの頭の動きによって追跡され、中心窩を中心とした画像を維持する。 目は広く間隔をあけられ、横に置かれ、近距離で広い双眼視野および精密な立体視をできます。 それはその頭を回転させるように移動する各目の暗いスポットは、偽瞳孔です。 これは、”正面に”見られるommatidiaが入射光を吸収し、側面にあるものがそれを反射するために発生します。
彼らの狩猟は視力に大きく依存しているので、カマキリは主に日中です。 多くの種は、しかし、夜に飛ぶし、人工の光に魅了されることがあります。 夜間に収集されたLiturgusidae科のカマキリは、主に男性であることが示されています。 夜行性の飛行は彼らのフェロモンの検出によってより少なく移動性の女性を見つけることの男性にとって特に重要です。 夜間の飛行は、昼間の飛行よりも少ない鳥の捕食者にカマキリを公開します。 多くのカマキリにも聴覚胸部器官があり、エコーロケーション呼び出しを検出して回避的に応答することによってコウモリを避けるのに役立ちます。
ダイエットと狩猟編集

カマキリは節足動物の一般的な捕食者である。 カマキリの大部分は、彼らの手の届くところに生きている獲物にのみ餌を与える待ち伏せ捕食者です。 彼らは自分自身をカモフラージュし、静止したまま、獲物が近づくのを待っているか、ゆっくりとしたステルスの動きで獲物を茎ます。 より大きなカマキリは、時にはトカゲ、カエル、魚、特に小さな鳥のような小さな脊椎動物だけでなく、自分の種のより小さな個体を食べる。
ほとんどのカマキリは、それが十分に近くに迷っている場合、魅力的な獲物を茎、彼らは特に空腹であるときにさらに行くでしょう。
手の届くところに入ると、カマキリはスパイクされた猛禽類の前足で獲物をつかむために急速に打ちます。 いくつかの地面と樹皮の種は、より積極的な方法で獲物を追求しています。 例えば、地上カマキリ、Entella、Ligaria、Ligariellaなどのいくつかの属のメンバーは、虎のカブトムシがそうであるように、獲物を求めて乾燥した地面の上を走ります。
いくつかの種の前腸は昆虫の全長を伸ばし、後で消化のために獲物を貯蔵するために使用することができます。
いくつかの種の前腸は昆虫の長さ これは、断続的に摂食する昆虫において有利であり得る。 中国のカマキリは、より長く生き、より速く成長し、花粉を食べることができるときにより多くの若い生産します。
アンチプレデーター adaptationsEdit
カマキリは、カエル、トカゲ、鳥などの脊椎動物と、クモ、大型種のスズメバチ、アリなどの無脊椎動物によって捕食されています。 いくつかの種のタキテスのようないくつかの狩猟スズメバチはまた、彼らの若者を養うためにカマキリのいくつかの種を麻痺させます。 一般的に、カマキリはカモフラージュによって身を守り、ほとんどの種は捕食者を避け、獲物をよりよくスネアするために、葉や他の背景に似ているように不可解に着色されている。 裸地や樹皮などの均一に着色された表面に生息するものは、その存在を明らかにするかもしれない影を排除するために、背側に平坦化されている。 花のカマキリと呼ばれる別の家族からの種は積極的な模倣である:彼らは花粉や蜜を収集するために来る獲物を引き付けるのに十分説得力のある アフリカやオーストラリアのいくつかの種は、乾季の終わりに向かって脱皮した後に黒くなることができます。

カマキリは、スティック昆虫のように、昆虫がリズミカルで反復的な左右の動きをするロッキング行動を示す。 この挙動のために提案された機能には,風中を移動する植生との類似性による陰窩の増強が含まれる。 しかし、反復的な揺れの動きは、昆虫がそれらの相対的な動きによって背景から物体を識別することを可能にする上で最も重要であり、より単純な視 これらの一般的に座りがちな昆虫による揺動の動きは、視野内の物体の相対運動の源として飛行または走行を置き換える可能性がある。 アリはカマキリの捕食者である可能性があるため、Loxomantis、Orthodera、Statiliaなどの属は、他の多くの節足動物と同様に、それらを攻撃することを避けます。 この行動を利用して、いくつかの初期のカマキリを含む節足動物の様々な、彼らの捕食者を回避するためにアリを模倣します。
-
葉の擬態:Choeradodisは葉のような前翅と広がった緑の胸郭を持っています。
-
大人の女性の虹彩オラトリオは、前足と翼が広がり、口が開いた状態で飼育し、はったりの脅威の表示を行います。
-
宝石で飾られた花のカマキリ、Creobroter gemmatus:鮮やかな色の翼は、捕食者を驚かせるためにdeimaticディスプレイで突然開かれています。
-
いくつかのカマキリのニンフは捕食者を避けるためにアリを模倣します。
繁殖と生活史編集
温帯気候での交尾期は、典型的には秋に行われ、熱帯地域では、交配は年中いつでも発生する可能性があります。 求愛に続いて交尾するために、男性は通常、女性の背中に飛び乗って、彼の前足で彼女の胸郭と翼の基盤を握りしめます。 彼はそれから女性の腹部の先端の近くの特別な部屋で精液を沈殿させ、貯えるために彼の腹部をアーチ状にします。 雌は種に応じて10-400個の卵を産む。 卵は、典型的には、腹部の腺によって大量に生産される泡の中に沈着する。 この泡は硬化し、卵の塊と一緒にオテカと呼ばれる保護カプセルを作ります。 種に応じて、オテカは平らな表面に付着したり、植物の周りに包まれたり、地面に堆積したりすることさえできます。 卵の多様性と耐久性にもかかわらず、彼らはしばしば、特にいくつかの種の寄生虫スズメバチによって捕食されています。 いくつかの種では、主にTarachodidae科の地面と樹皮のカマキリでは、母親は卵を守ります。 不可解なTarachodes maurusは彼女の腹部が彼女の卵のカプセルをカバーして樹皮に身を置き、獲物を渡して待ち伏せし、卵が孵化するまでほとんど動いていません。 アメリカ合衆国南部のブルナーのスティックカマキリによって珍しい生殖戦略が採用されており、この種では雄は発見されておらず、雌は単為生殖的に繁殖する。 単為生殖によって再現する能力は、少なくとも二つの他の種、Sphodromantis viridisとMiomantis spで記録されています。 これらの種は通常性的に再現されますが。 温暖な気候では、成虫は冬には生き残れず、卵は休眠し、春に孵化します。
超目Dictyopteraの密接に関連する昆虫群のように、カマキリは三つのライフステージを通過します:卵、ニンフ、および大人(カマキリは半代謝性昆虫の一つです)。 より小さい種のために、卵はより大きい種のための4-6週に対して3-4週に孵化するかもしれません。 幼虫は成虫とは異なる色をしていることがあり、初期段階はしばしばアリの模倣である。 それはその外骨格を脱皮としてカマキリのニンフが大きく成長します。 脱皮は、種に応じて、成体の段階に達する前に5〜10回発生する可能性があります。 最終的な脱皮の後、ほとんどの種は翼を持っていますが、いくつかの種は、特に女性の性別では、翼のないまたは短翼(”短翼”)のままです。 カマキリの寿命は種によって異なり、小さいものは4-8週間生きることができ、大きな種は4-6ヶ月生きることができます。
-
Mantis religiosa mating (brown male, green female)
-
Stagmomantis carolina laying ootheca
-
Recently laid M. religiosa ootheca
-
Hatching from the ootheca
-
Sphodromantis lineola molting
Sexual cannibalismEdit

Sexual cannibalism is common among most predatory species of mantises in captivity. それは時々、男性と女性の出会いの約四分の一が女性によって食べられている男性につながる自然の集団で観察されています。 カマキリの捕食種の約90%が性的な共食いを示しています。 成人男性は通常、最初は女性を上回っていますが、女性が選択的に小さい男性を食べるため、その数は成人段階の後半でかなり同等である可能性があ Tenodera sinensisでは、男性の83%が女性との出会いの後に共食いを逃れるが、複数の交配が起こるので、男性が食べられる確率は累積的に増加する。
雌は(通常の獲物と同様に)雄の頭を噛んで摂食を開始することがあり、交尾が始まった場合、雄の動きは精子の送達においてさらに活発になる 初期の研究者は、交尾運動は頭部ではなく腹部の神経節によって制御されるため、男性の頭部の除去は、栄養を得ながら受精を強化するための女性の生殖戦略であると考えていた。 その後、この行動は侵入的な実験室観察のアーティファクトであるように見えました。 行動が現場で自然であるかどうか、または人間の観察者によって引き起こされる気晴らしの結果であるかどうかは議論の余地がある。 カマキリは非常に視覚的な生物であり、明るい光や動いている科学者など、実験室や現場での妨害に気づきます。 アドリブを与えられていた中国のカマキリ(彼らは空腹ではなかったように)実際に乱されていないままにしたときに精巧な求愛行動を表示しました。 男性は求愛ダンスで女性を係合し、彼女の関心を摂食から交配に変える。 このような状況下で、女性は彼女の前足の内側に着色されたeyespotsを点滅させることにより、防御的なdeimaticディスプレイで応答することが知られています。実験は、貧しい食生活の女性が良い食生活の女性よりも性的な共食いに従事する可能性が高いことを示しています。
性的な共食いの理由は議論されています。
性的な共食いの理由は議論されています。
いくつかは、従順な男性が子孫を生産することによって選択的な利点を得るという仮説を立てています; これは、共食いされている男性の交尾の持続時間の定量化可能な増加によって支持され、場合によっては受精の持続時間と機会の両方を倍増させる。 これは、男性が空腹の女性にもっと注意して近づくことが見られ、空腹の女性に長時間装着されたままであることが示された研究によって対比され、積極的に共食いを避ける男性が複数の女性と交尾する可能性があることを示している。 同じ研究はまた、空腹の女性は一般的によく供給されたものよりも少ない男性を集めていることがわかりました。 交尾後に降りる行為は、現時点では女性が最も頻繁に仲間を共食いするため、男性にとって危険です。 装着期間の増加は、男性が彼女の仲間を共食いする可能性が高い空腹の女性をマウント解除するために適切な時間を待つことを示すように見えま 実験により,環境中の性比がカマキリの雄の交尾行動を決定し,これが女性の共食い傾向に影響を与え,精子競争仮説を支持することが明らかになった。 これはさらに、女性を降ろすことは、男性が共食いの影響を受けやすくすることができることを示唆している。
Leave a Reply