8.5: Reseptortyrosinkinaser (RTKs)
Reseptortyrosinkinaser medierer respons på et stort antall signaler, inkludert peptidhormoner som insulin og vekstfaktorer som epidermal vekstfaktor. Som GPCRs binder reseptortyrosinkinaser et signal, og sender meldingen videre gjennom en rekke intracellulære molekyler, hvorav den siste virker på målproteiner for å endre tilstanden til cellen.
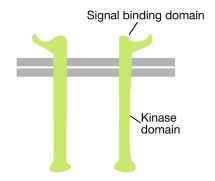
som navnet antyder, er en reseptortyrosinkinase en celleoverflatereseptor som også har en tyrosinkinaseaktivitet. Signalbindingsdomenet til reseptortyrosinkinasen er på celleoverflaten, mens tyrosinkinasens enzymatiske aktivitet ligger i den cytoplasmatiske delen av proteinet(se figur over). En transmembran alfa-helix forbinder disse to områdene av reseptoren.
Hva skjer når signalmolekyler binder seg til reseptortyrosinkinaser?
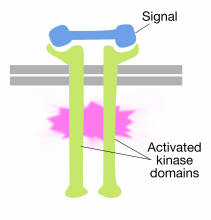
Binding av signalmolekyler til de ekstracellulære domenene av reseptor tyrosinkinasemolekyler fører til at to reseptormolekyler dimerer (kommer sammen og forbinder). Dette bringer cytoplasmatiske haler av reseptorene nær hverandre og forårsaker at tyrosinkinaseaktiviteten til disse haler blir slått på. De aktiverte haler fosforylerer deretter hverandre på flere tyrosinrester. Dette kalles autofosforylering.
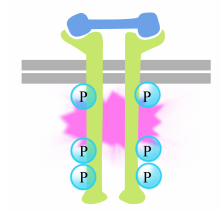
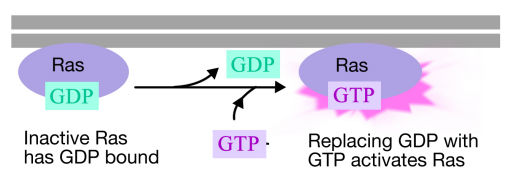
fosforyleringen av tyrosiner på reseptorhaler utløser sammenstillingen av en intracellulær signalkompleks på haler. De nylig fosforylerte tyrosinene tjener som bindingssteder for signalering av proteiner som deretter sender meldingen videre til andre proteiner. Et viktig protein som senere aktiveres av signalkompleksene på reseptortyrosinkinaser kalles Ras.Ras-proteinet er et monomert guaninnukleotidbindende protein som er forbundet med plasmamembranets cytosoliske ansikt (faktisk er det mye som alfa-underenheten til trimeriske g-proteiner). Akkurat som alfa – underenheten Til Et g-protein, er Ras aktiv når GTP er bundet til DET og inaktiv når BNP er bundet til DET.Også, som alfa-underenheten, Kan Ras hydrolysere GTP TIL BNP.
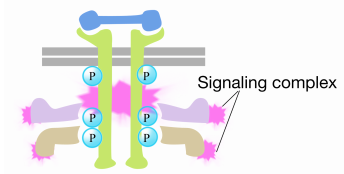
når et signal kommer til reseptortyrosinkinasen, kommer reseptormonomerer sammen og fosforylerer hverandres tyrosiner, og utløser samlingen av et kompleks av proteiner på den cytoplasmatiske halen av reseptoren. Et av proteinene i dette komplekset interagerer Med Ras og stimulerer utvekslingen AV BNP bundet til det inaktive Ras for EN GTP. Dette aktiverer Ras.
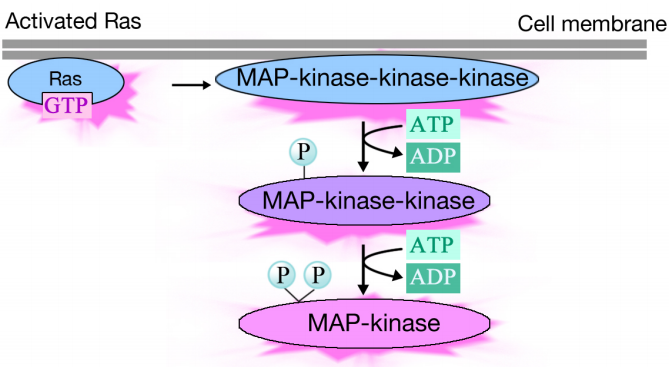
Aktivert Ras utløser en fosforyleringskaskade av tre proteinkinaser, som videresender og distribuerer signalet. DISSE proteinkinasene er medlemmer av EN gruppe KALT MAP-kinaser (Mitogenaktiverte Proteinkinaser). Den endelige kinasen i denne kaskade fosforylerer ulike målproteiner, inkludert enzymer og transkripsjonelle aktivatorer som regulerer genuttrykk.fosforyleringen av forskjellige enzymer kan endre deres aktiviteter og utløse nye kjemiske reaksjoner i cellen, mens fosforyleringen av transkripsjonelle aktivatorer kan endre hvilke gener som uttrykkes. Den kombinerte effekten av endringer i genuttrykk og proteinaktivitet endrer cellens fysiologiske tilstand.
Igjen, i følge banen til signaltransduksjon mediert Av Rtk, er det mulig å skjelne det samme grunnleggende mønster av hendelser: et signal er bundet av de ekstracellulære domenene til reseptortyrosinkinaser, noe som resulterer i reseptordimerisering og autofosforylering av cytosoliske haler, og formidler dermed meldingen til det indre av cellen.
meldingen sendes videre via et signalkompleks Til Ras som deretter stimulerer en rekke kinaser. Den terminale kinasen i kaskaden virker på målproteiner og forårsaker endringer i proteinaktiviteter og genuttrykk.
beskrivelsene ovenfor gir en veldig enkel skisse av noen av de store klassene av reseptorer og omhandler primært de mekanistiske detaljene i trinnene der signaler mottatt av ulike typer reseptorer gir endringer i celler. En stor take-home leksjon er den essensielle likheten til de forskjellige banene.Et annet poeng å huske på er at mens vi har sett på hver enkelt vei i isolasjon, mottar en celle til enhver tid flere signaler som utløser en rekke forskjellige svar samtidig. Banene beskrevet ovenfor viser en betydelig grad av «cross-talk» og responsen på et gitt signal påvirkes av de andre signalene som cellen mottar samtidig. Mangfoldet av forskjellige reseptorer, signaler og kombinasjonene derav er de midler som celler er i stand til å reagere på et enormt utvalg av forskjellige forhold.
Bidragsytere
-
Dr. Kevin Ahern og Dr. Indira Rajagopal (Oregon State University)
Leave a Reply