Hva Påvirker Størrelsen På Grupper Der Primater Velger Å Leve?

En av de store teoretiske bidrag gjort ved å studere primater er en forståelse av hva som påvirker naturen av komplekse variasjon i sosial struktur og organisasjoner. Underliggende dette er et sett av teoretiske utviklinger som undersøker hvorfor dyr lever i grupper i utgangspunktet og hvorfor grupper er av en bestemt størrelse. Her fokuserer vi på spørsmålet om «hva påvirker størrelsen på grupper primater velger å leve i?»Primater er spesielt nyttige taxa for å løse dette spørsmålet fordi deres gruppestørrelser er svært variable, ikke bare innenfor, men også mellom arter. Intraspesifikk gruppestørrelse for rød colobus (Procolobus rufomitratus) varierer for eksempel mellom 12 og 150 medlemmer (Chapman & Chapman 2000a). Interspesifikk variasjon varierer fra et enkelt individ, sett i orangutanger (Pongo sp.) (van Schaik 1999), til over 800 medlemmer i mandrills (Mandrillus sphinx) (Abernethy et al. 2002). Videre innenfor noen arter, sosiale grupper gjentatte ganger dele og re-forene i undergrupper av forskjellige størrelser og kombinasjoner over tid (f. eks, fisjon-fusion sosial organisering av edderkopp aper(Ateles sp. det finnes også en rekke andre typer sjimpanser og sjimpanser (Pan troglodytes), eller multi-level organisasjon av gelada (Theropithecus gelada) og Hamadryas bavianer (Papio hamadryas, Aureli et al. 2008). Denne variasjonen gir grunnlag for at forskere kan utvikle modeller for å undersøke både økologiske og sosiale drivere av gruppestørrelse.
Gruppering er gunstig på flere måter. Personer i større grupper antas å ha redusert risiko for predasjon (Hamilton 1971), kan være bedre i stand til å finne og forsvare matressurser (Cody 1971, Wrangham 1980), og kan være beskyttet mot spesifikk trussel, som infanticide av ekstra gruppe menn (Wrangham 1979). Ulike forskere har antydet at gruppering gir slike forutsigbare fordeler (Alexander 1974, van Schaik 1983)at forskjeller i gruppestørrelse kan forklares av ulempene (Wrangham et al. 1993). Den mest aksepterte potensielle kostnaden for gruppering antas å være en reduksjon i foraging effektivitet. Å være sammen med andre individer med samme diettbehov betyr at dyr enten kjemper om mat (konkurranse konkurranse), eller ett dyr i en gruppe slår en annen til maten, og når det andre dyret kommer til et område, er det rett og slett ingen mat igjen (scramble competition, Janson & van Schaik 1988). I begge disse situasjonene antas det at konkurranse om mat fører til at dyr må reise lenger. Logikken bak dette argumentet er relativt enkel. Dyr må beite over et område som kan møte deres energiske og ernæringsmessige krav. Det følger at en økning i gruppestørrelse vil øke arealet som må dekkes for å finne tilstrekkelige matforsyninger. Dermed må enkeltpersoner reise videre og bruke mer energi hvis de er i en stor gruppe, enn om de spiser i en mindre gruppe. Med en økning i reisetiden nærmer seg et punkt hvor energien som tilbys i reisen er for kostbar og mindre grupper blir fordelaktige. På denne måten kan økologiske faktorer påvirke bevegelsesmønstre og foraging effektivitet, og dermed begrense størrelsen på grupper som effektivt kan utnytte tilgjengelige matressurser. Disse ideene har blitt formalisert i Det Som har blitt kjent Som Den Økologiske Begrensninger Modell (Chapman & Chapman 2000b, Ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
den vesentlige komponenten i den økologiske begrensningsmodellen er at en økning i gruppestørrelse må føre til en økning i innen-gruppe matkonkurranse. Det er tenkelig at dette opererer på en litt annen måte, avhengig av arten av ressursene som brukes av bestemte arter. Med frugivorøse, og muligens mange folivorøse primater, som spiser i diskrete flekker-typisk trær som bærer matvarer eller klumper av trær-kan flere gruppemedlemmer tømme flekker raskere og føre til økte dagområder (Chapman 1988, Snaith & Chapman 2005). For flere insektsaktige arter, hvis ressurser ikke kan forekomme i så diskrete flekker, kan kontinuerlig reise gjennom baldakinen på jakt etter insekter med flere gruppemedlemmer føre til en økning i overlappingen av individuelle søkefelt, redusere antall møter per innbygger med mat og dermed øke området som må søkes (van Schaik et al. 1983).
så den økologiske begrensningsmodellen antyder at de faktorene som påvirker avstanden som dyr reiser, også bør påvirke gruppestørrelsen. For de dyrene som vanligvis spiser på frukt eller blader som kan tømmes, vil størrelsen på lappen avgjøre hvor lenge en gruppe av en gitt størrelse kunne bli og mate. En stor gruppe vil bruke mindre tid i en patch av en gitt størrelse enn en mindre gruppe, fordi den tømmer lappen raskere – en stor gruppe har bare flere munner å mate. Hvis dyr reiser mellom patcher når de har tømt dem, vil tettheten og fordelingen av patcher bestemme reisekostnadene som påløper. Når ressursoppdateringer er i høy tetthet eller i klumpfordeling, er avstanden til neste patch liten, reisekostnadene er lave, og dyr kan derfor danne store grupper. På slike tidspunkter kan eventuelle ekstra kostnader forbundet med å være medlem av en stor gruppe, for eksempel behovet for å besøke mange patcher, lett gjenopprettes. I kontrast, når ressursoppdateringer forekommer ved lave tettheter, er avstanden til neste patch vanligvis stor, reisekostnadene er høye, og dyr har ikke råd til å raskt tømme flekker, og derfor danner dyr små grupper. fisjon-fusion sosial organisering av edderkoppaper, sjimpanser og noen andre primater (Figur 1) tilbyr nyttige tester av den økologiske begrensningsmodellen fordi man kan forsøke å forutsi tidsmessige endringer i undergruppestørrelse fra direkte tiltak av størrelse, tetthet og distribusjon av matressurser (Chapman 1990, Chapman et al. 1995). For edderkoppaper og sjimpanser ble det utviklet en multippel regresjonsmodell for å vise at størrelsen, tettheten og fordelingen av matplaster målt hver måned var en god prediktor for undergruppestørrelse. Det er vanskelig å visuelt representere en multippel regresjon, men metodene som ble brukt for edderkoppaper er portrettert I Figur 2. Spider aper har en svært fleksibel fisjon-fusion type sosial organisasjon, Og I Santa Rosa National Park, Costa Rica, undergruppe størrelse kan variere fra 1 til 35 personer, men i gjennomsnitt ca 5 personer er funnet reiser sammen. For denne populasjonen kan 50% av variansen i gjennomsnittlig månedlig undergruppestørrelse forutsies fra relativt grove tiltak av størrelse, tetthet og distribusjon av matplaster (Chapman 1990).

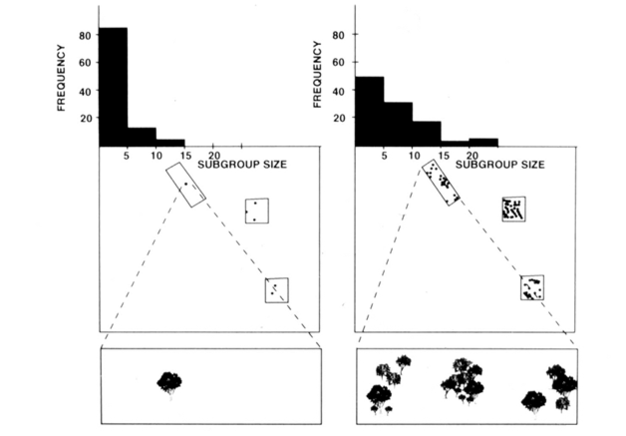
I tillegg til de ovennevnte artene har anvendelsen av den økologiske begrensningsmodellen generelt blitt støttet med forskning som kommer fra en rekke arter (Snaith & Chapman 2005) og situasjoner (f.eks. 1993, og blandede artsforeninger, Chapman & Chapman 2000c). Vi ser at, gitt den omfattende støtten som denne modellen har mottatt, er tiden kommet for at det er rimelig å foreslå at arter typisk skal overholde forventningene til den økologiske begrensningsmodellen. Dette betyr ikke at alle arter vil samsvare, og dermed blir det en spennende tid for forskning på dette feltet fordi etterforskere kan søke etter unntak fra modellen. Vi mistenker at arter som ikke samsvarer vil gjøre det fordi de har vedtatt sosiale strategier som strider mot de økologiske forventningene. Dette gir en «målestokk» for å evaluere den potensielle betydningen av ulike sosiale strategier(dvs. jo lenger en avviker fra forventningene til den økologiske modellen, desto viktigere er det sosiale selektive presset).
la oss kort gi to eksempler fra vår tidligere forskning. Kvinnelige sjimpanser var mer ensomme enn kvinnelige edderkoppaper, men de har begge lignende fisjonsfusjon sosiale organisasjoner. Videre, mens vi nøyaktig kunne forutsi antall menn og subadult sjimpanser i en undergruppe basert på økologiske forhold, var vi ikke i stand til å forutsi antall kvinnelige sjimpanser i disse undergruppene (Chapman et al. 1995). Selv når ressursene var ekstremt rikelig og nesten alle mennene var i en stor undergruppe, gikk kvinner sjelden inn i grupper, noe som tyder på at kostnaden for å være i en større undergruppe oppveide noen fordeler, for eksempel rovdyrunngåelse. Dette er uventet fra det perspektivet at spedbarnene til disse kvinnene ville være alder / kjønnsklasse mest truet av predasjon. En testbar hypotese for å forklare disse observasjonene er at naturen til koalisjonene i disse to artene kan påvirke fordelene med gruppemedlemskap. Bevis tyder på at, i motsetning til sjimpanser, danner edderkoppaper kvinner koalisjoner som ofte opererer for å tillate medlemmer av koalisjonen eksklusiv tilgang til mat. Så edderkoppaper, når de blir med i bestemte personer, kan øke tilgangen til mat ved å ekskludere andre, men for sjimpansehunner er dette ikke tilfelle (Chapman et al. 1995). Vi identifiserte en annen interessant avvik, potensielt forårsaket av sosiale faktorer som påvirker kostnadene og fordelene ved å være i en gruppe, og dette gjelder røde colobus og svart-hvite colobusaber (aka. guerezas, Colobus guereza) ved Kibale Nasjonalpark I Uganda. Rød colobus danner store grupper med et gjennomsnitt på 65 personer (25-127 personer, Snaith et al. 2008), mens guerezaer lever i små grupper med et gjennomsnitt på 6,5 individer (4-11 individer, Harris & Chapman 2007). Til tross for denne forskjellen er det stor likhet i plantene spist av de to artene. Hvis økologiske forhold var ansvarlige for forskjellen i gruppestørrelse mellom de to colobinartene, ville man forvente at deres dietter ville variere og tettheten av mattrær ville være lavere i guerezas hjemområder, siden de har den mindre gruppestørrelsen. Men deres dietter er svært like; kostholdsoverlappingen mellom en rød colobus og en guereza-gruppe som hadde sitt hjemområde helt innenfor hjemmeområdet til den røde colobus-gruppen var 43.2%, mens for de to nabogruppene av rød colobus, overlappet diett med bare 37,3% (Chapman et al. 2002). Det ser ut til at kvinnelig guereza reproduktiv suksess er maksimert i små og mellomstore grupper, enten fordi større grupper opplever høyere forekomst av overtak og barnedrap eller mer matkonkurranse (se bevis på scramble konkurranse i folivorous Trachypithecus phayrei og Semnopithecus sp. , men mangel på bevis på guerezas tappe mat patcher ). Vi har sett en rekke mannlige overtakelser og spedbarnsdrap i guerezaene (Onderdonk 2000) — se Teichroeb & Sicotte (2008) for en evaluering av teori om barnedrap i kolobiner. Det som ikke er klart er hvordan risikoen for barnedrap vil begrense størrelsen på guereza-grupper, når de har tilsynelatende tilgang til en høy tetthet av mat som bør tillate dannelsen av multi-mannlige grupper. I flere arter er større hunngrupper mer av et mål for innvandrende menn, og denne situasjonen kan øke barnedrapsraten-for Eksempel Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). Barnedrap forventes imidlertid å være mindre hyppig hvis en gruppe inneholder flere reproduktivt aktive menn, fordi mannlig innvandring og påfølgende voldelig overtakelse og barnedrap er mindre sannsynlig å oppstå når nykommeren står overfor flere mulige sirer av fremtidige spedbarn (Janson & van Schaik 2000). Guerezas synes å ha rikelig matressurser til å danne større grupper (Tombak et al. 2012), som ville redusere risikoen for barnedrap, men de danner ikke store grupper. Det er mulig at naturen av mannlige-mannlige interaksjoner i guerezas gjør store grupper ustabile. Dette ser ut til å være tilfelle i ursine colobus (Colobus vellerosus), hvor multi-mannlige grupper tiltrekker mannlige innvandrere og har høyere spedbarnsdrap enn grupper med en sterk voksen mann i sin prime (Teichroeb, Wikberg, Badescu, & Sicotte, upubliserte data; Figur 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

Hvis modellen for økologiske begrensninger beskriver trykk som generelt påvirker primatgruppestørrelsen, og hvis avvik fra denne forventede gruppestørrelsen skyldes sosialt press, vil det være en spennende tid på dette forskningsområdet fordi det reiser en rekke interessante spørsmål. Eksempelvis: Hva er arten av det sosiale presset på gruppestørrelsen? Hvorfor er disse sosiale begrensningene tilstede i disse artene og ikke nært beslektede? Og hva er de evolusjonære kreftene som velger for disse sosiale pressene? Gruppestørrelse er et kritisk element som påvirker typen sosial organisasjon arter vedta, og dermed undersøke disse spørsmålene vil føre til spennende forbedringer av sosioekologiske modeller.
Leave a Reply