8.5: Receptortyrosinekinasen (RTKs)
Receptortyrosinekinasen mediëren reacties op een groot aantal signalen, waaronder peptidehormonen zoals insuline en groeifactoren zoals epidermale groeifactor. Als GPCRs, binden de kinases van de receptortyrosine een signaal, dan geven het bericht door een reeks intracellular molecules door, waarvan de laatste op doelproteã nen handelt om de staat van de cel te veranderen.
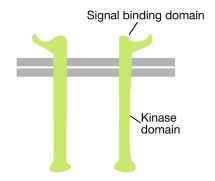
zoals de naam al doet vermoeden, is een receptortyrosinekinase een celoppervlak-receptor die ook een tyrosinekinase-activiteit heeft. Het signaalbindingsdomein van het receptortyrosinekinase bevindt zich op het celoppervlak, terwijl de enzymatische activiteit van het tyrosinekinase zich bevindt in het cytoplasmatische deel van het eiwit (zie figuur hierboven). Een transmembrane alpha helix verbindt deze twee gebieden van de receptor.
Wat gebeurt er als signaalmoleculen zich binden aan receptortyrosinekinasen?
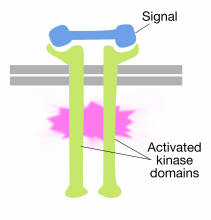
Binding van signaalmoleculen aan de extracellulaire domeinen van receptortyrosinekinasemoleculen veroorzaakt dimerisatie van twee receptormoleculen (kom samen en associëren). Dit brengt de cytoplasmic staarten van de receptoren dicht bij elkaar en veroorzaakt de tyrosinekinase activiteit van deze staarten worden aangezet. De geactiveerde staarten fosforyleren elkaar op verschillende tyrosineresiduen. Dit wordt autofosforylatie genoemd.
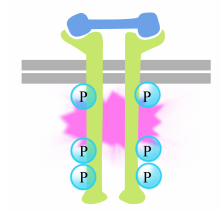
de fosforylering van tyrosines op de receptorstaarten veroorzaakt de assemblage van een intracellulair signaalcomplex op de staarten. De pas phosphorylated tyrosines dienen als bandplaatsen voor signalerende proteã nen die dan het bericht aan nog andere proteã nen doorgeven. Een belangrijke proteã ne die later door de signalerende complexen op de receptortyrosinekinases wordt geactiveerd wordt genoemd Ras.
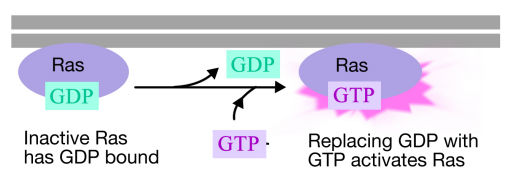
het Ras-eiwit is een monomeer guaninenucleotidebindend eiwit dat geassocieerd is met het cytosolisch gelaat van het plasmamembraan (in feite lijkt het sterk op de Alfa-subeenheid van trimere G-eiwitten). Net als de Alfa – subeenheid van een g-eiwit is Ras actief wanneer GTP eraan gebonden is en inactief wanneer GDP eraan gebonden is.Ook kan Ras, net als de Alfa-subeenheid, de GTP hydrolyseren tot het BBP.
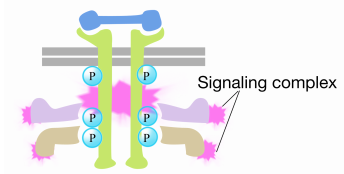
wanneer een signaal aankomt bij het receptortyrosinekinase, komen de receptormonomeren samen en fosforylaat elkaars tyrosines, waardoor de assemblage van een complex van eiwitten op de cytoplasmatische staart van de receptor wordt geactiveerd. Één van de proteã nen in dit complexe interageert met Ras en bevordert de uitwisseling van het BBP dat aan het inactieve Ras voor GTP wordt gebonden. Dit activeert het Ras.
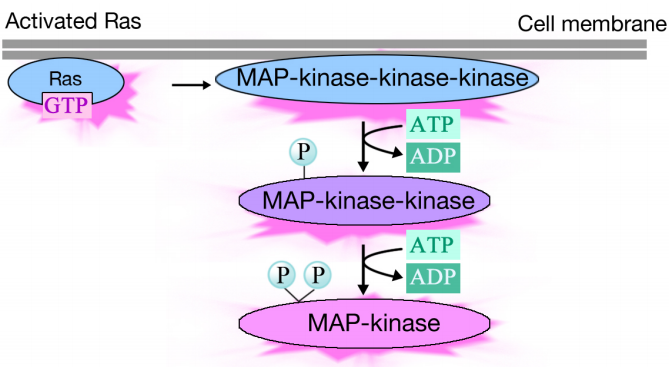
geactiveerd Ras activeert een fosforyleringscascade van drie eiwitkinasen, die het signaal relais en distribueren. Deze eiwitkinasen zijn leden van een groep genoemd de KAARTKINASEN (Mitogen geactiveerde eiwitkinasen). Het definitieve kinase in deze cascade phosphorylates diverse doelproteã nen, met inbegrip van enzymen en transcriptional activators die genuitdrukking regelen.
de fosforylering van verschillende enzymen kan hun activiteiten veranderen en nieuwe chemische reacties in de cel veroorzaken, terwijl de fosforylering van transcriptionele activatoren kan veranderen welke genen worden uitgedrukt. Het gecombineerde effect van veranderingen in genuitdrukking en eiwitactiviteit verandert de fysiologische toestand van de cel.
bij het volgen van het pad van signaaltransductie gemedieerd door RTKs, is het mogelijk om hetzelfde basispatroon van gebeurtenissen te onderscheiden: een signaal wordt gebonden door de extracellulaire domeinen van receptortyrosinekinasen, resulterend in receptordimerisatie en autofosforylatie van de cytosolic staarten, waarbij de boodschap aan het binnenland van de cel wordt overgebracht.
het bericht wordt via een signaleringscomplex doorgegeven aan Ras, dat vervolgens een reeks kinasen stimuleert. Het terminale kinase in de cascade handelt op doelproteã nen en brengt in veranderingen in eiwitactiviteiten en genuitdrukking over.
de bovenstaande beschrijvingen geven een zeer eenvoudige schets van enkele van de belangrijkste klassen van receptoren en behandelen voornamelijk de mechanistische details van de stappen waarmee signalen die door verschillende soorten receptoren worden ontvangen veranderingen in cellen teweegbrengen. Een belangrijke take-home Les is de essentiële gelijkenis van de verschillende routes.
een ander punt om in gedachten te houden is dat terwijl we elk individueel traject afzonderlijk hebben bekeken, een cel op een bepaald moment meerdere signalen ontvangt die een verscheidenheid aan verschillende reacties tegelijk veroorzaken. De hierboven beschreven wegen tonen een aanzienlijke mate van “cross-talk” en de reactie op om het even welk gegeven signaal wordt beà nvloed door de andere signalen die de cel gelijktijdig ontvangt. De veelheid aan verschillende receptoren, signalen en de combinaties daarvan zijn de middelen waarmee cellen kunnen reageren op een enorme verscheidenheid aan verschillende omstandigheden.
bijdragers
-
Dr. Kevin Ahern en Dr. Indira Rajagopal (Oregon State University)
Leave a Reply