Wat beïnvloedt de grootte van groepen waarin primaten kiezen om te leven?

een van de belangrijkste theoretische bijdragen van het bestuderen van primaten is een inzicht in wat de aard van complexe variatie in sociale structuur en organisaties beïnvloedt. Aan de basis hiervan ligt een reeks theoretische ontwikkelingen die onderzoeken waarom dieren in de eerste plaats in groepen leven en waarom groepen van een bepaalde grootte zijn. Hier richten we ons op de vraag “wat beïnvloedt de grootte van groepen waarin primaten kiezen om te leven?”
primaten zijn bijzonder nuttige taxa om deze vraag te beantwoorden, omdat hun groepsgrootte zeer variabel is, niet alleen binnen, maar ook tussen soorten. De grootte van de intraspecifieke Groep voor rode colobus (Procolobus rufomitratus) varieert bijvoorbeeld tussen 12 en 150 leden (Chapman & Chapman 2000a). Interspecifieke variatie varieert van een enkel individu, zoals gezien in orang-oetans (Pongo sp.) (van Schaik 1999), aan meer dan 800 leden in mandrills (Mandrillus sphinx) (Abernethy et al. 2002). Bovendien, binnen sommige soorten, sociale groepen herhaaldelijk te verdelen en opnieuw te verenigen in subgroepen van verschillende grootte en combinaties in de tijd (bijvoorbeeld, kernsplijting-fusion sociale organisatie van spin apen (Ateles sp.) en chimpansees (Pan troglodytes), of de multi-level organisatie van gelada (Theropithecus gelada) en hamadryas bavianen (Papio hamadryas, Aureli et al. 2008). Deze variatie biedt de basis voor onderzoekers om modellen te ontwikkelen om zowel de ecologische als de sociale drijfveren van groepsgrootte te onderzoeken.
groepering is op verschillende manieren nuttig. Individuen in grotere groepen worden verondersteld een verminderd risico op predatie te hebben( Hamilton 1971), kunnen beter in staat zijn om voedselbronnen te vinden en te verdedigen (Cody 1971, Wrangham 1980), en kunnen worden beschermd tegen conspecifieke dreiging, zoals kindermoord door extra-groepsmannetjes (Wrangham 1979). Verschillende onderzoekers hebben gesuggereerd dat groepering zulke voorspelbare voordelen biedt (Alexander 1974, van Schaik 1983) dat verschillen in groepsgrootte kunnen worden verklaard door de nadelen (Wrangham et al. 1993). De meest algemeen aanvaarde potentiële kosten van groepering worden verondersteld om een vermindering van de foerageerefficiëntie te zijn. Samen zijn met andere individuen met dezelfde voedingsbehoeften betekent dat dieren ofwel vechten om voedsel (wedstrijd competitie), of een dier in een groep verslaat een ander op het voedsel, dus wanneer het tweede dier in een gebied komt is er gewoon geen voedsel meer over (scramble competitie, Janson & van Schaik 1988). In beide situaties wordt gedacht dat de concurrentie om voedsel ertoe leidt dat dieren verder moeten reizen. De logica achter dit argument is relatief eenvoudig. Dieren moeten foerageren over een gebied dat aan hun energetische en voedingsbehoeften kan voldoen. Het gevolg is dat een toename van de groepsgrootte zal leiden tot een toename van het gebied dat moet worden bestreken om voldoende voedselvoorraden te vinden. Zo moeten individuen verder reizen en meer energie uitgeven als ze in een grote groep zitten, dan als ze in een kleinere groep foerageren. Met een toename van de tijd besteed aan reizen, wordt een punt benaderd waar de energie die wordt besteed aan reizen te duur is en kleinere groepen voordelig worden. Op deze manier kunnen ecologische factoren bewegingspatronen en foerageerefficiëntie beïnvloeden, waardoor de grootte van groepen die de beschikbare voedselbronnen efficiënt kunnen benutten, wordt beperkt. Deze ideeën zijn geformaliseerd in wat bekend is geworden als het Ecological Constraints Model (Chapman & Chapman 2000b, Ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
de essentiële component van het model voor ecologische beperkingen is dat een toename van de groepsgrootte moet leiden tot een toename van de binnen-groep voedende concurrentie. Het is denkbaar dat dit op een iets andere manier werkt, afhankelijk van de aard van de hulpbronnen die door bepaalde soorten worden gebruikt. Met fruitetende, en mogelijk veel bladetende primaten, die zich voeden in discrete plekken – meestal bomen met voedsel of klontjes bomen-kunnen extra groepsleden vlekken sneller afbreken en leiden tot een groter aantal dagen (Chapman 1988, Snaith & Chapman 2005). Voor meer insectenetende soorten, waarvan de middelen niet kunnen voorkomen in als discrete patches, kan continue reizen door het bladerdak op zoek naar insecten met extra groepsleden leiden tot een toename van de overlapping van individuele zoekvelden, het verminderen per hoofd van de bevolking tegenkomen tarieven met voedsel en dus het vergroten van het gebied dat moet worden gezocht (Van Schaik et al. 1983).
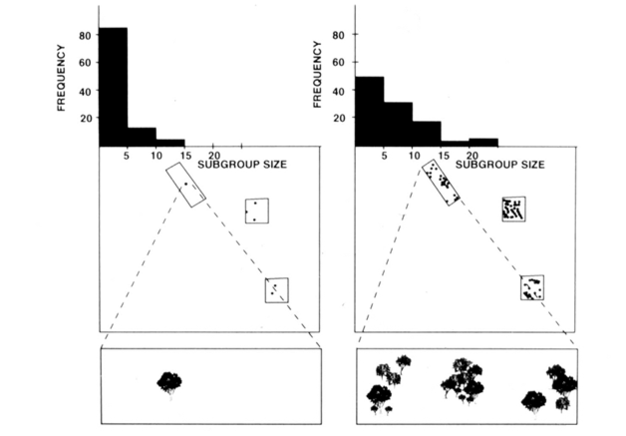
het ecological constraints model suggereert dus dat de factoren die van invloed zijn op de afstand die dieren afleggen, ook van invloed moeten zijn op de groepsgrootte. Voor dieren die zich gewoonlijk voeden met vruchten of bladeren die uitgeput kunnen zijn, zou de grootte van de patch bepalen hoe lang een groep van een bepaalde grootte zou kunnen blijven en voeden. Een grote groep zou minder tijd doorbrengen in een patch van een bepaalde grootte dan een kleinere groep, omdat het put de patch sneller — een grote groep heeft gewoon meer monden te voeden. Als dieren tussen pleisters reizen nadat ze uitgeput zijn, dan zullen de dichtheid en verdeling van pleisters de gemaakte reiskosten bepalen. Wanneer resource patches zijn op een hoge dichtheid of in een cluped distributie, de afstand tot de volgende patch is klein, de reiskosten zijn laag, en dieren kunnen daarom grote groepen vormen. Op dergelijke momenten kunnen eventuele extra kosten in verband met het lidmaatschap van een grote groep, zoals de noodzaak om vele patches te bezoeken, gemakkelijk worden hersteld. In tegenstelling, wanneer de hulpbronflarden bij lage dichtheden voorkomen, is de afstand tot de volgende flard typisch groot, zijn de reiskosten hoog, en kunnen de dieren zich niet veroorloven om flarden snel uit te putten, en daarom vormen de dieren kleine groepen. de fission-fusion social organization of spider monkeys, chimpansees, and a few other primates (Fig.1) biedt nuttige tests van het ecological constraints model omdat men kan proberen temporele veranderingen in de grootte van subgroepen te voorspellen aan de hand van directe metingen van de grootte, dichtheid en distributie van voedselbronnen (Chapman 1990, Chapman et al. 1995). Voor spinapen en chimpansees werd een multiple regression model ontwikkeld om aan te tonen dat de grootte, dichtheid en distributie van voedselpleisters die elke maand worden gemeten, een goede voorspeller waren van de grootte van de subgroep. Het is moeilijk om visueel een meervoudige regressie weer te geven, maar de methoden die werden gebruikt voor spinnenapen worden afgebeeld in Figuur 2. Spider monkeys hebben een zeer flexibele kernsplijting-fusion soort sociale organisatie, en in Santa Rosa National Park, Costa Rica, subgroep grootte kan variëren van 1 tot 35 individuen, maar gemiddeld ongeveer 5 individuen worden gevonden reizen samen. Voor deze populatie kan 50% van de variantie in gemiddelde maandelijkse subgroepgrootte worden voorspeld aan de hand van relatief ruwe metingen van de grootte, dichtheid en distributie van voedselpleisters (Chapman 1990).

naast de bovengenoemde soorten is de toepasbaarheid van het ecological constraints model over het algemeen ondersteund met onderzoek afkomstig van een verscheidenheid aan soorten (Snaith & Chapman 2005) en situaties (bijv. general models, Wrangham et al. 1993, and mixed species associations, Chapman & Chapman 2000c). Wij zijn van mening dat, gezien de brede steun die dit model heeft gekregen, de tijd is gekomen dat het redelijk is om te suggereren dat soorten typisch moeten voldoen aan de verwachtingen van het model van ecologische beperkingen. Dit betekent niet dat alle soorten zich zullen conformeren, en dus wordt het een spannende tijd voor onderzoek op dit gebied, omdat onderzoekers kunnen zoeken naar uitzonderingen op het model. Wij vermoeden dat soorten die zich niet conformeren dat zullen doen omdat zij sociale strategieën hebben aangenomen die tegen de ecologische verwachtingen ingaan. Dit verschaft een “maatstaf” om het potentiële belang van verschillende sociale strategieën te evalueren (d.w.z. hoe verder men afwijkt van de verwachtingen van het ecologische model, hoe belangrijker de sociale selectieve druk).
laten we kort twee voorbeelden geven uit ons vorige onderzoek. Vrouwelijke chimpansees waren meer solitair dan vrouwelijke spinnenapen, maar ze hebben allebei vergelijkbare sociale organisaties voor kernsplijting. Verder, hoewel we het aantal mannetjes en subadult chimpansees in een subgroep nauwkeurig konden voorspellen op basis van ecologische omstandigheden, waren we niet in staat om het aantal vrouwelijke chimpansees in deze subgroepen te voorspellen (Chapman et al. 1995). Zelfs toen de middelen zeer overvloedig waren en bijna alle mannetjes in één grote subgroep zaten, gingen de vrouwtjes zelden in groepen, wat suggereert dat de kosten om in een grotere subgroep te zitten opwegen tegen eventuele voordelen, zoals het vermijden van roofdieren. Dit is onverwacht vanuit het perspectief dat de zuigelingen van deze wijfjes de leeftijd/geslachtsklasse zouden zijn die het meest door predatie worden bedreigd. Een testbare hypothese om deze waarnemingen te verklaren is dat de aard van de coalities in deze twee soorten de voordelen van groepslidmaatschap kan beïnvloeden. Er zijn aanwijzingen dat, in tegenstelling tot chimpansees, spinnenaapjes coalities vormen die vaak opereren om de leden van de coalitie exclusieve toegang te geven tot voedsel. Dus spinnenapen, wanneer ze zich bij bepaalde individuen voegen, kunnen hun toegang tot voedsel vergroten door anderen uit te sluiten, maar voor chimpansee-vrouwen is dit niet het geval (Chapman et al. 1995).
we identificeerden een andere interessante afwijking, mogelijk veroorzaakt door sociale factoren die van invloed zijn op de kosten en baten van een groep, en dit betreft rode colobus en zwart-witte colobusapen (aka. guerezas, Colobus guereza) in Kibale National Park, Oeganda. Rode colobus vormen grote groepen met een gemiddelde van 65 individuen (25-127 individuen, Snaith et al. Guerezas leven in kleine groepen met gemiddeld 6,5 individuen (4-11 individuen, Harris & Chapman 2007). Ondanks dit verschil is er een grote overeenkomst in de planten gegeten door de twee soorten. Als de ecologische omstandigheden verantwoordelijk waren voor het verschil in groepsgrootte tussen de twee colobines, zou men verwachten dat hun diëten zouden verschillen en dat de dichtheid van voedselbomen lager zou zijn in het thuisbereik van de guerezas, omdat ze de kleinere groepsgrootte hebben. Hun diëten zijn echter zeer vergelijkbaar; de overlap tussen een rode colobus en een guereza-groep die zijn thuisbereik volledig binnen het thuisbereik van de rode colobus-groep had was 43.2%, terwijl voor de twee naburige groepen van rode colobus, dieet overlapte met slechts 37,3% (Chapman et al . 2002). Het lijkt erop dat vrouwelijke guereza reproductief succes wordt gemaximaliseerd in kleine en middelgrote groepen, hetzij omdat grotere groepen ervaren hogere percentages van take-overs en infanticide of meer voeding competitie (zie bewijs van scramble competitie in de folivore Trachypithecus phayrei en Semnopithecus sp. , maar een gebrek aan bewijs van guerezas uitputting van voedsel patches). We zijn getuige geweest van een aantal mannelijke overnames en kindermoorden in de guerezas (Onderdonk 2000)-zie Teichroeb & Sicotte (2008) voor een evaluatie van de theorie met betrekking tot infanticide in colobines. Wat niet duidelijk is, is hoe het risico van infanticide de grootte van guereza-groepen zou beperken, wanneer zij duidelijk toegang hebben tot een hoge dichtheid van voedsel dat de vorming van multi-mannelijke groepen mogelijk zou moeten maken. In verschillende soorten, Grotere vrouwelijke groepen zijn meer van een doelwit voor immigrerende mannetjes, en deze situatie kan de kindermoord tarieven verhogen – bijvoorbeeld, Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). De verwachting is echter dat kindermoord minder vaak voorkomt als een groep meerdere reproductief actieve mannen bevat, omdat mannelijke Immigratie en daaropvolgende gewelddadige overname en kindermoord minder waarschijnlijk zijn wanneer de nieuwkomer wordt geconfronteerd met verschillende mogelijke nakomelingen van toekomstige zuigelingen (Janson & van Schaik 2000). Guerezas lijken voldoende voedselbronnen te hebben om grotere groepen te vormen (Tombak et al. 2012), waardoor het risico op kindermoord zou afnemen, maar ze vormen geen grote groepen. Het is mogelijk dat de aard van man-man interacties in guerezas grote groepen onstabiel maakt. Dit blijkt het geval te zijn bij ursine colobus (Colobus vellerosus), waar multi-mannelijke groepen mannelijke immigranten aantrekken en hogere kindermoordpercentages hebben dan groepen met één sterke volwassen man in zijn bloei (Teichroeb, Wikberg, Badescu, & Sicotte, niet-gepubliceerde gegevens; Figuur 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

indien het ecological constraints model een overzicht geeft van de druk die in het algemeen van invloed is op de grootte van de primatengroep en indien afwijkingen van deze verwachte groepsgrootte het gevolg zijn van sociale druk, zal het een spannende tijd zijn op dit gebied van onderzoek omdat het een aantal interessante vragen oproept. Bijvoorbeeld: Wat is de aard van de sociale druk op de groepsgrootte? Waarom zijn deze sociale beperkingen aanwezig in deze soorten en niet nauw verwante? En wat selecteren de evolutionaire krachten voor deze sociale druk? Groepsgrootte is een cruciaal element dat van invloed is op het type sociale organisatie dat soorten aannemen, dus het onderzoeken van deze kwesties zal leiden tot spannende verfijningen van socio-ecologische modellen.
Leave a Reply