8.5: Receptortyrosinkinaser (RTK)
Receptortyrosinkinaser förmedlar svar på ett stort antal signaler, inklusive peptidhormoner som insulin och tillväxtfaktorer som epidermal tillväxtfaktor. Liksom GPCRs binder receptortyrosinkinaser en signal och skickar sedan meddelandet vidare genom en serie intracellulära molekyler, varav den sista verkar på målproteiner för att förändra cellens tillstånd.
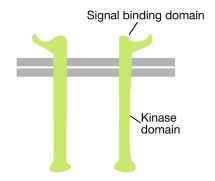
som namnet antyder är ett receptortyrosinkinas en cellytreceptor som också har en tyrosinkinasaktivitet. Signalbindningsdomänen för receptortyrosinkinas är på cellytan, medan tyrosinkinasenzymaktiviteten finns i den cytoplasmiska delen av proteinet (se figur ovan). En transmembran alfa-helix förbinder dessa två regioner i receptorn.
vad händer när signalmolekyler binder till receptortyrosinkinaser?
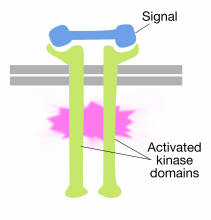
bindning av signalmolekyler till receptorns extracellulära domäner tyrosinkinasmolekyler får två receptormolekyler att dimerisera (komma ihop och associera). Detta ger de cytoplasmiska svansarna i receptorerna nära varandra och gör att tyrosinkinasaktiviteten hos dessa svansar slås på. De aktiverade svansarna fosforylerar sedan varandra på flera tyrosinrester. Detta kallas autofosforylering.
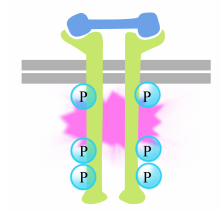
fosforyleringen av tyrosiner på receptorns svansar utlöser sammansättningen av en intracellulärt signalkomplex på svansarna. De nyligen fosforylerade tyrosinerna fungerar som bindningsställen för signalering av proteiner som sedan skickar meddelandet vidare till ännu andra proteiner. Ett viktigt protein som därefter aktiveras av signalkomplexen på receptortyrosinkinaserna kallas Ras.
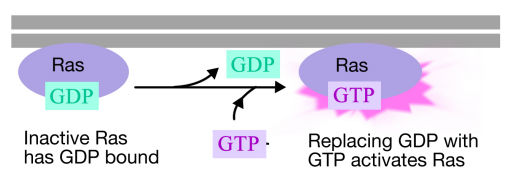
Ras-proteinet är ett monomeriskt guaninnukleotidbindande protein som är associerat med plasmamembranets cytosoliska ansikte (det är faktiskt mycket som alfa-subenheten av trimera G-proteiner). Precis som alfa – subenheten för ett g-protein är Ras aktiv när GTP är bunden till det och inaktiv när BNP är bunden till det.Liksom alfa-subenheten kan Ras hydrolysera GTP till BNP.
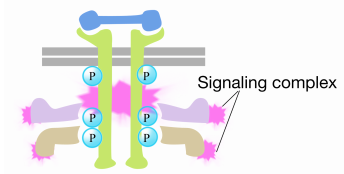
När en signal anländer till receptortyrosinkinas, samlas receptormonomererna och fosforylerar varandras tyrosiner, vilket utlöser sammansättningen av ett komplex av proteiner på receptorns cytoplasmiska svans. Ett av proteinerna i detta komplex interagerar med Ras och stimulerar utbytet av BNP bunden till den inaktiva Ras för en GTP. Detta aktiverar Ras.
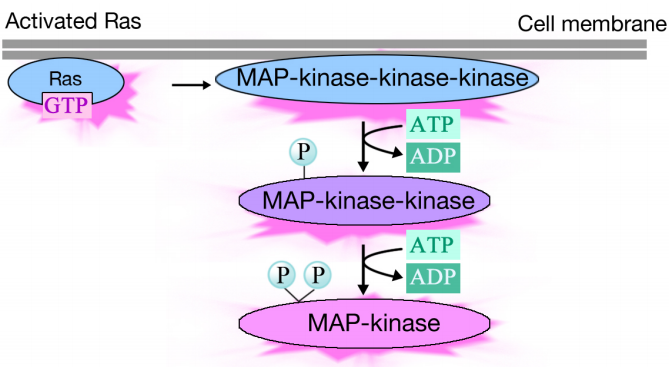
aktiverad Ras utlöser en fosforyleringskaskad av tre proteinkinaser, som reläer och distribuerar signalen. Dessa proteinkinaser är medlemmar i en grupp som kallas MAP-kinaser (Mitogenaktiverade proteinkinaser). Det slutliga kinaset i denna kaskadfosforylerar olika målproteiner, inklusive enzymer och transkriptionella aktivatorer som reglerar genuttryck.
fosforyleringen av olika enzymer kan förändra deras aktiviteter och utlösa nya kemiska reaktioner i cellen, medan fosforyleringen av transkriptionella aktivatorer kan ändra vilka gener som uttrycks. Den kombinerade effekten av förändringar i genuttryck och proteinaktivitet förändrar cellens fysiologiska tillstånd.
återigen, genom att följa signaltransduktionsvägen medierad av RTK, är det möjligt att urskilja samma grundläggande mönster av händelser: en signal är bunden av de extracellulära domänerna av receptortyrosinkinaser, vilket resulterar i receptordimerisering och autofosforylering av de cytosoliska svansarna, vilket förmedlar meddelandet till cellens inre.
meddelandet vidarebefordras via ett signalkomplex till Ras som sedan stimulerar en serie kinaser. Terminalkinas i kaskaden verkar på målproteiner och medför förändringar i proteinaktiviteter och genuttryck.
beskrivningarna ovan ger en mycket enkel skiss av några av de stora klasserna av receptorer och handlar främst om de mekanistiska detaljerna i stegen genom vilka signaler som mottas av olika typer av receptorer medför förändringar i celler. En stor take-home lektion är den väsentliga likheten mellan de olika vägarna.
en annan punkt att tänka på är att medan vi har tittat på varje enskild väg isolerat, mottar en cell vid varje given tidpunkt flera signaler som utlöser en mängd olika svar på en gång. De vägar som beskrivs ovan visar en avsevärd grad av” korsprat ” och svaret på en given signal påverkas av de andra signalerna som cellen mottar samtidigt. Mängden olika receptorer, signaler och kombinationerna därav är det sätt på vilket celler kan svara på en enorm mängd olika omständigheter.
bidragsgivare
-
Dr Kevin Ahern och Dr Indira Rajagopal (Oregon State University)
Leave a Reply