Vad påverkar storleken på grupper där primater väljer att leva?

ett av de viktigaste teoretiska bidragen genom att studera primater är en förståelse för vad som påverkar naturen av komplex variation i social struktur och organisationer. Bakom detta ligger en uppsättning teoretiska utvecklingar som undersöker varför djur lever i grupper i första hand och varför grupper är av en viss storlek. Här, vi fokuserar på frågan om ” vad påverkar storleken på grupper primater väljer att leva i?”
primater är särskilt användbara taxa för att ta itu med denna fråga eftersom deras gruppstorlekar är mycket varierande, inte bara inom, men också mellan arter. Intraspecifik gruppstorlek för röd colobus (Procolobus rufomitratus) varierar till exempel mellan 12 och 150 medlemmar (Chapman & Chapman 2000a). Interspecifik variation varierar från en enskild individ, vilket ses i orangutanger (Pongo sp.) (van Schaik 1999), till över 800 medlemmar i mandrills (Mandrillus sphinx) (Abernethy et al. 2002). Inom vissa arter delar sig dessutom sociala grupper upprepade gånger och återförenas i undergrupper av olika storlekar och kombinationer över tiden (t.ex. fission-fusion social organization of spider monkeys (Ateles sp.) och schimpanser (Pan troglodytes), eller organisationen på flera nivåer av gelada (Theropithecus gelada) och Hamadryas babianer (Papio hamadryas, Aureli et al. 2008). Denna variation ger grunden för forskare att utveckla modeller för att undersöka både ekologiska och sociala drivkrafter för gruppstorlek.
gruppering är fördelaktig på flera sätt. Individer i större grupper tros ha en minskad risk för predation (Hamilton 1971), kan bättre kunna hitta och försvara livsmedelsresurser (Cody 1971, Wrangham 1980) och kan skyddas mot specifikt hot, som barnmord av extragruppsmän (Wrangham 1979). Olika forskare har föreslagit att gruppering ger sådana förutsägbara fördelar (Alexander 1974, van Schaik 1983) att skillnader i gruppstorlek kan förklaras av nackdelarna (Wrangham et al. 1993). Den mest accepterade potentiella kostnaden för gruppering anses vara en minskning av födosökningseffektiviteten. Att vara med andra individer med samma kostbehov innebär att djur antingen kämpar om mat (tävlingstävling) eller ett djur i en grupp slår ett annat till maten, så när det andra djuret kommer till ett område finns det helt enkelt ingen mat kvar (scramble competition, Janson & van Schaik 1988). I båda dessa situationer tros det att konkurrens om mat leder till att djur måste resa längre. Logiken bakom detta argument är relativt enkel. Djur måste foder över ett område som kan uppfylla deras energiska och näringsmässiga krav. Av detta följer att en ökning av gruppstorleken kommer att öka det område som måste täckas för att hitta tillräckliga livsmedel. Således måste individer resa längre och spendera mer energi om de är i en stor grupp, än om de foder i en mindre grupp. Med en ökning av tiden som reser, närmar sig en punkt där energin som spenderas i resor är för dyr och mindre grupper blir fördelaktiga. På detta sätt kan ekologiska faktorer påverka rörelsemönster och födosökningseffektivitet, vilket begränsar storleken på grupper som effektivt kan utnyttja tillgängliga livsmedelsresurser. Dessa tankar har formaliserats i vad som har blivit känt som den ekologiska Begränsningsmodellen (Chapman & Chapman 2000b, ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
den väsentliga komponenten i modellen för ekologiska begränsningar är att en ökning av gruppstorleken måste leda till en ökning av foderkonkurrensen inom gruppen. Det är tänkbart att detta fungerar på ett något annorlunda sätt beroende på arten av de resurser som används av vissa arter. Med frugivorösa och eventuellt många folivorösa primater, som matar i diskreta fläckar-vanligtvis träd som bär matvaror eller klumpar av träd — kan ytterligare gruppmedlemmar tömma lappar snabbare och leda till ökade dagintervall (Chapman 1988, Snaith & Chapman 2005). För mer insektsätande arter, vars resurser kanske inte förekommer i så diskreta fläckar, kan kontinuerlig resa genom baldakinen på jakt efter insekter med ytterligare gruppmedlemmar leda till en ökning av överlappningen av enskilda sökfält, vilket minskar antalet möten per capita med mat och därmed ökar det område som måste sökas (van Schaik et al. 1983).
så den ekologiska begränsningsmodellen antyder att de faktorer som påverkar avståndet som djur reser också bör påverka gruppstorleken. För de djur som vanligtvis matar på frukt eller löv som kan tömmas, skulle plåstrets storlek avgöra hur länge en grupp av en viss storlek kunde stanna och mata. En stor grupp skulle spendera mindre tid i en lapp av en viss storlek än en mindre grupp, eftersom den tömmer plåstret snabbare — en stor grupp har helt enkelt fler munnar att mata. Om djur reser mellan lappar när de har tömt dem, kommer tätheten och fördelningen av lappar att avgöra resekostnaderna. När resurspatchar har hög densitet eller i en klumpig fördelning är avståndet till nästa lapp litet, resekostnaderna är låga och djur kan därför bilda stora grupper. Vid sådana tillfällen kan eventuella extra kostnader i samband med att vara medlem i en stor grupp, såsom behovet av att besöka många patchar, lätt återvinnas. Däremot, när resurspatchar uppträder vid låga densiteter, är avståndet till nästa lapp vanligtvis stort, resekostnaderna är höga och djur har inte råd att snabbt tömma lappar och därför bildar djur små grupper.
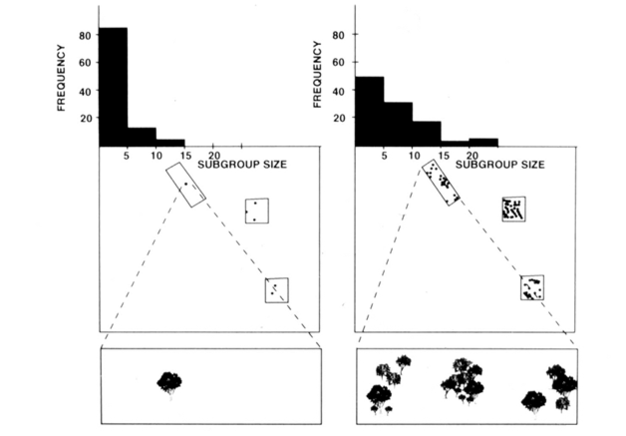
fission-fusion social organisation av spindelapor, schimpanser och några andra primater (Figur 1) erbjuder användbara tester av den ekologiska begränsningsmodellen eftersom man kan försöka förutsäga tidsmässiga förändringar i undergruppsstorlek från direkta mått på storlek, densitet och fördelning av livsmedelsresurser (Chapman 1990, Chapman et al. 1995). För spindelapor och schimpanser utvecklades en multipel regressionsmodell för att visa att storleken, densiteten och fördelningen av matplåster som mättes varje månad var en bra prediktor för undergruppsstorlek. Det är svårt att visuellt representera en multipel regression, men metoderna som användes för spindelapor visas i Figur 2. Spindelapor har en mycket flexibel fission-fusion typ av social organisation, och i Santa Rosa National Park, Costa Rica, kan undergruppsstorlek variera från 1 till 35 individer, men i genomsnitt finns cirka 5 individer som reser tillsammans. För denna befolkning kan 50% av variansen i genomsnittlig månadsstorlek undergrupp förutsägas från relativt råa mått på storlek, densitet och fördelning av matplåster (Chapman 1990).

förutom de arter som nämns ovan har tillämpligheten av den ekologiska begränsningsmodellen generellt stödts med forskning från en mängd olika arter (Snaith & Chapman 2005) och situationer (t.ex. allmänna modeller, Wrangham et al. 1993, och blandade artsföreningar, Chapman & Chapman 2000c). Vi anser att med tanke på det omfattande stöd som denna modell har fått är det dags att det är rimligt att föreslå att arter vanligtvis ska överensstämma med förväntningarna på modellen för ekologiska begränsningar. Detta betyder inte att alla arter kommer att överensstämma, och därmed blir det en spännande tid för forskning inom detta område eftersom utredare kan söka efter undantag från modellen. Vi misstänker att arter som inte överensstämmer kommer att göra det eftersom de har antagit sociala strategier som strider mot de ekologiska förväntningarna. Detta ger en” måttstock ” för att utvärdera den potentiella betydelsen av olika sociala strategier (dvs. ju längre man avviker från förväntningarna på den ekologiska modellen, desto viktigare är det sociala selektiva trycket).
låt oss kortfattat ge två exempel från vår tidigare forskning. Kvinnliga schimpanser var mer ensamma än kvinnliga spindelapor, men de har båda liknande fission-fusion sociala organisationer. Vidare kunde vi exakt förutsäga antalet män och subadulta schimpanser i en undergrupp baserad på ekologiska förhållanden, men vi kunde inte förutsäga antalet kvinnliga schimpanser i dessa undergrupper (Chapman et al. 1995). Även när resurserna var extremt rikliga och nästan alla män var i en stor undergrupp, gick kvinnor sällan in i grupper, vilket tyder på att kostnaden för att vara i en större undergrupp uppvägde några fördelar, såsom undvikande av rovdjur. Detta är oväntat ur perspektivet att dessa kvinnors spädbarn skulle vara den ålder/könsklass som mest hotas av predation. En testbar hypotes för att förklara dessa observationer är att arten av koalitionerna i dessa två arter kan påverka fördelarna med gruppmedlemskap. Bevis tyder på att, till skillnad från schimpanser, spider monkey-kvinnor bildar koalitioner som ofta fungerar för att tillåta medlemmarna i koalitionen exklusiv tillgång till mat. Så spindelapor, när de går med i vissa individer, kan öka sin tillgång till mat genom att utesluta andra, men för schimpanskvinnor är detta inte fallet (Chapman et al. 1995).
vi identifierade en annan intressant avvikelse, potentiellt orsakad av sociala faktorer som påverkar kostnaderna och fördelarna med att vara i en grupp, och detta gäller röd colobus och svartvita colobus apor (aka. Guerezas, Colobus guereza) vid Kibale nationalpark, Uganda. Röd colobus bildar stora grupper med i genomsnitt 65 individer (25-127 individer, Snaith et al. 2008), medan guerezas bor i små grupper med i genomsnitt 6,5 individer (4-11 individer, Harris & Chapman 2007). Trots denna skillnad finns det stor likhet i de växter som äts av de två arterna. Om ekologiska förhållanden var ansvariga för skillnaden i gruppstorlek mellan de två colobine-arterna, skulle man förvänta sig att deras dieter skulle skilja sig och densiteten hos matträd skulle vara lägre i guerezas hemområden, eftersom de har den mindre gruppstorleken. Men deras dieter är mycket lika; kostöverlappningen mellan en röd colobus och en guereza-grupp som hade sitt hemområde helt inom den röda colobus-gruppens hemområde var 43.2%, medan de två angränsande grupperna av röd colobus överlappade diet med endast 37,3% (Chapman et al. 2002). Det verkar som om kvinnlig guereza reproduktiv framgång maximeras i små och medelstora grupper, antingen för att större grupper upplever högre take-overs och barnmord eller mer utfodringstävling (se bevis på rusningstävling i folivorous Trachypithecus phayrei och Semnopithecus sp. , men brist på bevis på att guerezas tömmer matplåster ). Vi har bevittnat ett antal manliga take-overs och spädbarnsmord i guerezas (Onderdonk 2000) — se Teichroeb & Sicotte (2008) för en utvärdering av teorin om barnmord i colobines. Det som inte är klart är hur risken för barnmord skulle begränsa storleken på guereza-grupper, när de har uppenbar tillgång till en hög densitet av mat som bör möjliggöra bildandet av flerhanliga grupper. I flera arter är större kvinnliga grupper mer ett mål för invandrande män, och denna situation kan öka barnamordspriserna-till exempel Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). Barnmord förväntas dock vara mindre frekvent om en grupp innehåller flera reproduktivt aktiva män, eftersom manlig invandring och efterföljande våldsamt övertagande och barnmord är mindre benägna att inträffa när nykomlingen står inför flera möjliga sires av framtida spädbarn (Janson & van Schaik 2000). Guerezas verkar ha gott om matresurser för att bilda större grupper (Tombak et al. 2012), vilket skulle minska risken för barnmord, men de bildar inte stora grupper. Det är möjligt att karaktären hos manliga-manliga interaktioner i guerezas gör stora grupper instabila. Detta verkar vara fallet i ursine colobus (Colobus vellerosus), där flera manliga grupper lockar manliga invandrare och har högre barnamord än grupper med en stark vuxen man i sin främsta (Teichroeb, Wikberg, Badescu, & Sicotte, opublicerade data; Figur 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

om modellen för ekologiska begränsningar beskriver tryck som generellt påverkar primatgruppens storlek och om avvikelser från denna förväntade gruppstorlek beror på socialt tryck kommer det att bli en spännande tid inom detta forskningsområde eftersom det väcker ett antal intressanta frågor. Exempelvis: Vad är det sociala trycket på gruppstorlek? Varför finns dessa sociala begränsningar i dessa arter och inte nära besläktade? Och vad väljer de evolutionära krafterna för dessa sociala tryck? Gruppstorlek är ett kritiskt element som påverkar typen av social organisation arter anta, således undersöka dessa frågor kommer att leda till spännande förbättringar av socioekologiska modeller.
Leave a Reply