Expresiones moleculares Biología celular: Endosomas
Endosomas y endocitosis
Los endosomas son vesículas unidas a membrana, formadas a través de una familia compleja de procesos conocidos colectivamente como endocitosis, y que se encuentran en el citoplasma de prácticamente todas las células animales. El mecanismo básico de la endocitosis es el inverso de lo que ocurre durante la exocitosis o la secreción celular. Implica la invaginación (plegamiento hacia adentro) de la membrana plasmática de una célula para rodear macromoléculas u otra materia que se difunde a través del fluido extracelular. Los materiales extraños rodeados se introducen en la célula y, después de un pellizco de la membrana (denominado brote), se liberan al citoplasma en una vesícula en forma de saco. El tamaño de las vesículas varía, y las de más de 100 nanómetros de diámetro se conocen típicamente como vacuolas.
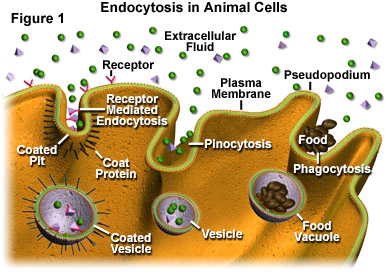
En la Figura 1 se ilustran tres mecanismos primarios de endocitosis que se exhiben en una célula típica. En el extremo izquierdo de la figura, se presenta la endocitosis mediada por receptores, que es la forma más específica del proceso endocítico. A través de la endocitosis mediada por receptores, las células activas son capaces de absorber cantidades significativas de moléculas particulares (ligandos) que se unen a los sitios receptores que se extienden desde la membrana citoplasmática hasta el fluido extracelular que rodea a la célula. Estos sitios receptores se agrupan comúnmente a lo largo de fosas recubiertas en la membrana, que están revestidas en su superficie citoplasmática con una colección de proteínas en forma de cerdas. Se cree que las proteínas de la capa juegan un papel en la ampliación de la fosa y la formación de una vesícula. Tenga en cuenta que, como se muestra en la Figura 1, las vesículas producidas a través de la endocitosis mediada por receptores pueden internalizar otras moléculas además de los ligandos, aunque los ligandos generalmente se introducen en la célula en una concentración más alta.
Un mecanismo menos específico de endocitosis es la pinocitosis, que se ilustra en la sección central de la Figura 1. Por medio de la pinocitosis, una célula es capaz de ingerir gotitas de líquido del fluido extracelular. Todos los solutos que se encuentran en las gotitas fuera de la célula pueden quedar encerrados en las vesículas formadas a través de este proceso, y los presentes en la mayor concentración en el fluido extracelular también se vuelven los más concentrados en los sacos membranosos. Las vesículas pinocíticas tienden a ser más pequeñas que las producidas por otros procesos endocíticos.
El tipo final de endocitosis, denominado fagocitosis (véase la Figura 1), es probablemente la forma más conocida en que una célula puede importar materiales externos. En muchos laboratorios de ciencias escolares, los niños observan amebas bajo el microscopio y observan a los organismos unicelulares comer estirando pseudopodios y rodeando cualquier partícula de alimento que encuentren en su camino. Este envolvimiento y posterior empaquetado de las partículas en vesículas, que generalmente son lo suficientemente grandes como para denominarse correctamente vacuolas, es fagocitosis. Aunque comúnmente se asocia con amebas, la fagocitosis es practicada por muchos organismos. En la mayoría de los animales multicelulares, las células fagocíticas funcionan principalmente en defensa corporal en lugar de como un medio para obtener alimento. Por ejemplo, los leucocitos en el cuerpo humano a menudo fagocitan protozoos, bacterias, células muertas y materiales similares para ayudar a evitar infecciones u otros problemas.
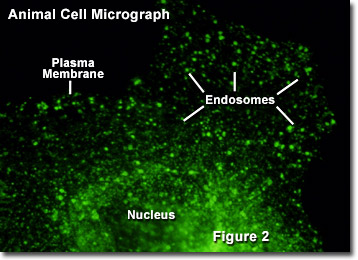
Una vez liberadas en el citoplasma, varias vesículas pequeñas producidas por endocitosis pueden unirse para formar una sola entidad. Este endosoma generalmente funciona de una de dos maneras. Lo más común es que los endosomas transporten su contenido en una serie de pasos a un lisosoma, que posteriormente digiere los materiales. En otros casos, sin embargo, los endosomas son utilizados por la célula para transportar varias sustancias entre diferentes porciones de la membrana celular externa. Esta última función es particularmente importante entre las células epiteliales, como las que componen la capa externa de la piel, porque exhiben polaridad (un lado de la célula es diferente del otro lado). En la Figura 2 se ilustra una imagen digital de fluorescencia de una única célula de fibroblastos de riñón de mono verde africano (línea CV-1) transfectada con una proteína fluorescente fusionada a una secuencia de aminoácidos dirigida para endosomas (verde). El núcleo, la membrana plasmática y los componentes del endosoma están marcados en la figura.
Un endosoma que está destinado a transferir su contenido a un lisosoma generalmente pasa por varios cambios a lo largo de su camino. En su forma inicial, cuando la estructura se conoce a menudo como un endosoma temprano, la vesícula especializada contiene un solo compartimento. Con el tiempo, sin embargo, se producen cambios químicos en la vesícula y la membrana que rodea el endosoma se pliega sobre sí misma de una manera similar a la invaginación de la membrana plasmática. En este caso, sin embargo, la membrana no se pellizca. En consecuencia, se forma una estructura con múltiples compartimentos, denominada endosoma multivesicular. El endosoma multivesicular es una estructura intermedia en la que se producen cambios químicos adicionales, incluida una caída significativa en el nivel de pH, a medida que la vesícula se convierte en un endosoma tardío.
Aunque los endosomas tardíos son capaces de descomponer muchas proteínas y grasas, se necesita un lisosoma para digerir completamente todos los materiales que contienen. Con frecuencia, el contenido de los endosomas tardíos se transporta a un lisosoma a través de la fusión (unión) de sus membranas. En algunas circunstancias, los endosomas tardíos son capaces de madurar en lisosomas a través de modificaciones químicas y estructurales adicionales, en cuyo caso la fusión no es necesaria para completar la digestión.
VOLVER A LA ESTRUCTURA CELULAR DE LOS ANIMALES
¿Preguntas o comentarios? Envíenos un correo electrónico.
© 1995-2019 por Michael W. Davidson y la Universidad Estatal de Florida. Todos los Derechos Reservados. No se pueden reproducir ni utilizar de ninguna manera imágenes, gráficos, software, scripts o applets sin el permiso de los titulares de los derechos de autor. El uso de este sitio web significa que acepta todos los Términos y Condiciones Legales establecidos por los propietarios.
Este sitio web es mantenido por nuestro
Graphics & Equipo de programación web
en colaboración con Optical Microscopy en el
National High Magnetic Field Laboratory.
Última modificación: viernes, 13 de noviembre de 2015 a las 01:18 PM
Recuento de accesos Desde el 26 de mayo de 2005: 220036
Microscopios proporcionados por:
![]()
![]()
Leave a Reply