8.5: Receptortyrosinkinaser (RTKs)
Receptortyrosinkinaser formidler reaktioner på et stort antal signaler, herunder peptidhormoner som insulin og vækstfaktorer som epidermal vækstfaktor. Ligesom GPCR ‘ erne binder receptortyrosinkinaser et signal og sender derefter meddelelsen videre gennem en række intracellulære molekyler, hvoraf den sidste virker på målproteiner for at ændre cellens tilstand.
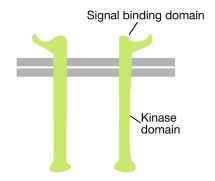
Som navnet antyder, er en receptortyrosinkinase en celleoverfladereceptor, der også har en tyrosinkinaseaktivitet. Det er en af de mest almindelige typer af protein, der findes i den cytoplasmatiske del af proteinet (se figur ovenfor). En transmembran alfa-spiral forbinder disse to regioner af receptoren.
Hvad sker der, når signalmolekyler binder til receptortyrosinkinaser?
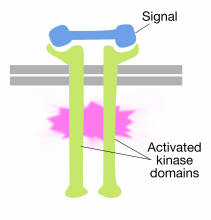
Binding af signalmolekyler til de ekstracellulære domæner af receptor tyrosinkinasemolekyler får to receptormolekyler til at dimerisere (komme sammen og associere). Dette bringer de cytoplasmatiske haler af receptorerne tæt på hinanden og får tyrosinkinaseaktiviteten af disse haler til at blive tændt. De aktiverede haler phosphorylerer derefter hinanden på flere tyrosinrester. Dette kaldes autophosphorylering.
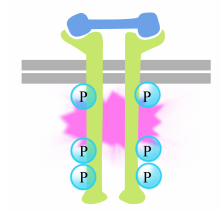
phosphoryleringen af tyrosin på receptorhalerne udløser samlingen af en intracellulært signalkompleks på halerne. De nyligt phosphorylerede tyrosiner tjener som bindingssteder til signalering af proteiner, der derefter videregiver budskabet til endnu andre proteiner. Et vigtigt protein, der efterfølgende aktiveres af signalkomplekserne på receptortyrosinkinaserne kaldes Ras.
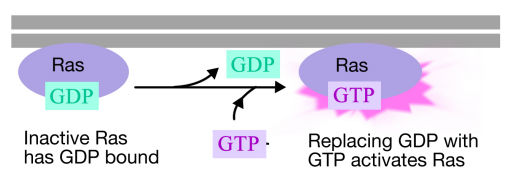
Ras-proteinet er et monomert guaninnukleotidbindende protein, der er forbundet med plasmamembranens cytosoliske ansigt (faktisk er det meget som alfa-underenheden af trimeriske G-proteiner). Ligesom alfa-underenheden af et G-protein er Ras aktiv, når GTP er bundet til det og inaktivt, når BNP er bundet til det.Ligesom alpha-underenheden kan Ras også hydrolysere GTP til BNP.
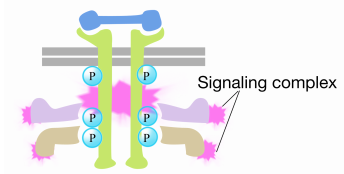
når et signal ankommer til receptortyrosinkinasen, kommer receptormonomererne sammen og phosphorylerer hinandens tyrosin, hvilket udløser samlingen af et kompleks af proteiner på receptorens cytoplasmatiske hale. Et af proteinerne i dette kompleks interagerer med Ras og stimulerer udvekslingen af BNP bundet til den inaktive Ras for en GTP. Dette aktiverer Ras.
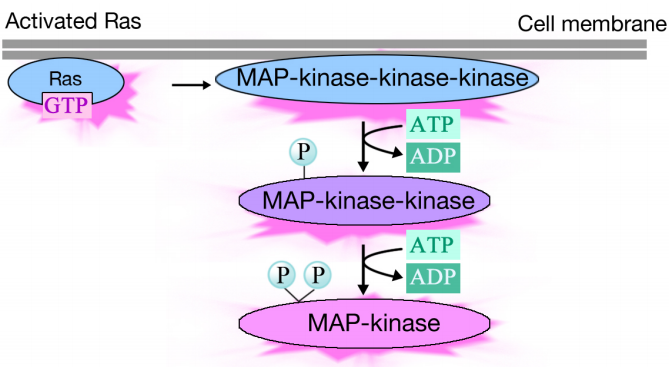
aktiveret Ras udløser en phosphoryleringskaskade af tre proteinkinaser, som relæ og distribuerer signalet. Disse proteinkinaser er medlemmer af en gruppe kaldet MAP-kinaser (MITOGENAKTIVEREDE proteinkinaser). Den endelige kinase i denne kaskade phosphorylerer forskellige målproteiner, herunder transkriptionsaktivatorer, der regulerer genekspression.fosforyleringen kan ændre deres aktiviteter og udløse nye kemiske reaktioner i cellen, mens phosphoryleringen af transkriptionsaktivatorer kan ændre, hvilke gener der udtrykkes. Den kombinerede effekt af ændringer i genekspression og proteinaktivitet ændrer cellens fysiologiske tilstand.
endnu en gang, ved at følge stien til signaltransduktion medieret af RTKs, er det muligt at skelne det samme grundlæggende mønster af begivenheder: et signal er bundet af de ekstracellulære domæner af receptortyrosinkinaser, hvilket resulterer i receptordimerisering og autophosphorylering af de cytosoliske haler, hvilket således formidler budskabet til det indre af cellen.
meddelelsen overføres via et signalkompleks til Ras, som derefter stimulerer en række kinaser. Den terminale kinase i kaskaden virker på målproteiner og medfører ændringer i proteinaktiviteter og genekspression.
beskrivelserne ovenfor giver en meget enkel skitse af nogle af de største klasser af receptorer og beskæftiger sig primært med de mekanistiske detaljer i de trin, hvormed signaler modtaget af forskellige typer receptorer medfører ændringer i celler. En stor take-home lektion er den væsentlige lighed mellem de forskellige veje.
et andet punkt at huske på er, at mens vi har set på hver enkelt vej isoleret, modtager en celle til enhver tid flere signaler, der udløser en række forskellige svar på en gang. De ovenfor beskrevne veje viser en betydelig grad af” cross-talk”, og responsen på et givet signal påvirkes af de andre signaler, som cellen modtager samtidigt. Mangfoldigheden af forskellige receptorer, signaler og kombinationerne deraf er de midler, hvormed celler er i stand til at reagere på en enorm række forskellige omstændigheder.
bidragydere
-
Dr. Kevin Ahern og Dr. Indira Rajagopal (Oregon State University)
Leave a Reply