hvad påvirker størrelsen af grupper, hvor primater vælger at leve?

et af de største teoretiske bidrag, der ydes ved at studere primater, er en forståelse af, hvad der påvirker arten af kompleks variation i social struktur og organisationer. Bag dette er et sæt teoretiske udviklinger, der undersøger, hvorfor dyr lever i grupper i første omgang, og hvorfor grupper er af en bestemt størrelse. Her fokuserer vi på spørgsmålet om “hvad påvirker størrelsen af grupper primater vælger at leve i?”
primater er særligt nyttige til at løse dette spørgsmål, fordi deres gruppestørrelser er meget variable, ikke kun inden for, men også mellem arter. Intraspecifik gruppestørrelse for rød colobus (Procolobus rufomitratus) varierer for eksempel mellem 12 og 150 medlemmer (Chapman & Chapman 2000A). Interspecifik variation spænder fra et enkelt individ, som det ses i orangutanger (Pongo sp.) (van Schaik 1999), til over 800 medlemmer i mandrills (Mandrillus Sfinks) (Abernethy et al. 2002). Desuden deler sociale grupper inden for nogle arter gentagne gange og forenes igen i undergrupper i forskellige størrelser og kombinationer over tid (f.eks. fission-fusion social organisation af edderkoppeaber (ateles sp.) og chimpanser (Pan troglodytes) eller organisationen på flere niveauer af gelada (Theropithecus gelada) og Hamadryas bavianer (Papio hamadryas, Aureli et al. 2008). Denne variation giver grundlaget for forskere til at udvikle modeller til at undersøge både de økologiske og de sociale drivkræfter i gruppestørrelse.
gruppering er gavnlig på flere måder. Personer i større grupper menes at have en nedsat risiko for rovdyr (Hamilton 1971), kan være bedre i stand til at finde og forsvare fødevareressourcer (Cody 1971, Krangham 1980) og kan beskyttes mod en specifik trussel, som barnemord af mænd uden for gruppen (Krangham 1979). Forskellige forskere har foreslået, at gruppering giver sådanne forudsigelige fordele (Aleksandr 1974, van Schaik 1983), at forskelle i gruppestørrelse kan forklares med ulemperne. 1993). De mest accepterede potentielle omkostninger ved gruppering menes at være en reduktion i fodereffektivitet. At være sammen med andre personer med de samme kostbehov betyder, at dyr enten kæmper om mad (konkurrence konkurrence), eller et dyr i en gruppe slår et andet til fødevaren, så når det andet dyr kommer til et område, er der simpelthen ingen mad tilbage (scramble konkurrence, Janson & van Schaik 1988). I begge disse situationer menes det, at konkurrence om mad fører til, at dyr skal rejse længere. Logikken bag dette argument er relativt enkel. Dyr skal foder over et område, der kan opfylde deres energiske og ernæringsmæssige krav. Det følger heraf, at en stigning i gruppestørrelse vil øge det område, der skal dækkes for at finde tilstrækkelige fødevareforsyninger. Således skal enkeltpersoner rejse længere og bruge mere energi, hvis de er i en stor gruppe, end hvis de foder i en mindre gruppe. Med en stigning i rejsetiden nærmer man sig et punkt, hvor den energi, der bruges i rejser, er for dyr, og mindre grupper bliver fordelagtige. På denne måde kan økologiske faktorer påvirke bevægelsesmønstre og fodereffektivitet og derved begrænse størrelsen på grupper, der effektivt kan udnytte tilgængelige fødevareressourcer. Disse ideer er blevet formaliseret i det, der er blevet kendt som den økologiske Begrænsningsmodel (Chapman & Chapman 2000B, Ganas & Robbins 2005, Snaith & Chapman 2007, Teichroeb & Sicotte 2009).
den væsentlige komponent i modellen for økologiske begrænsninger er, at en stigning i gruppestørrelse skal føre til en stigning i fodringskonkurrence inden for gruppen. Det kan tænkes, at dette fungerer på en lidt anden måde afhængigt af arten af de ressourcer, der anvendes af bestemte arter. Med sparsomme og muligvis mange folivorøse primater, der lever i diskrete pletter-typisk træer, der bærer madvarer eller klumper af træer — kan yderligere gruppemedlemmer nedbryde pletter hurtigere og føre til øgede dagintervaller (Chapman 1988, Snaith & Chapman 2005). For mere insektædende arter, hvis ressourcer muligvis ikke forekommer i som diskrete pletter, kan kontinuerlig rejse gennem baldakinen på jagt efter insekter med yderligere gruppemedlemmer føre til en stigning i overlapningen af individuelle søgefelter, hvilket reducerer antallet af møder pr.indbygger med mad og dermed øger det område, der skal søges (van Schaik et al. 1983).
så modellen for økologiske begrænsninger antyder, at de faktorer, der påvirker den afstand, som dyr rejser, også skal påvirke gruppestørrelsen. For de dyr, der typisk lever af frugt eller blade, der kan udtømmes, vil størrelsen på plasteret bestemme, hvor længe en gruppe af en given størrelse kunne blive og fodre. En stor gruppe ville bruge mindre tid i et plaster af en given størrelse end en mindre gruppe, fordi det udtømmer plasteret hurtigere — en stor gruppe har simpelthen flere mund at fodre. Hvis dyr rejser mellem patches, når de har udtømt dem, vil tætheden og fordelingen af patches bestemme de afholdte rejseomkostninger. Når ressourcepatcher har en høj densitet eller i en klumpet fordeling, er afstanden til den næste patch lille, rejseomkostningerne er lave, og dyr kan derfor danne store grupper. På sådanne tidspunkter kan eventuelle ekstraomkostninger forbundet med at være medlem af en stor gruppe, såsom behovet for at besøge mange programrettelser, let gendannes. I modsætning hertil, når ressourcepatcher forekommer ved lave tætheder, er afstanden til den næste patch typisk stor, rejseomkostningerne er høje, og dyr har ikke råd til hurtigt at nedbryde patches, og derfor danner dyr små grupper.
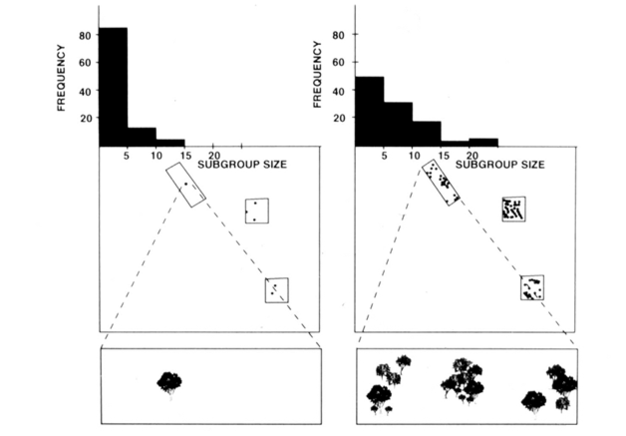
fission-fusion social organisation af edderkoppeaber, chimpanser og et par andre primater (Figur 1) tilbyder nyttige tests af den økologiske begrænsningsmodel, fordi man kan forsøge at forudsige tidsmæssige ændringer i undergruppestørrelse fra direkte målinger af størrelsen, densiteten og fordelingen af fødevareressourcer (Chapman 1990, Chapman et al. 1995). For edderkoppeaber og chimpanser blev der udviklet en multipel regressionsmodel for at vise, at størrelsen, densiteten og fordelingen af madplaster målt hver måned var en god forudsigelse for undergruppestørrelse. Det er svært at visuelt repræsentere en multipel regression, men de metoder, der blev brugt til edderkopaber, er afbildet i figur 2. Spider aber har en meget fleksibel fission-fusion type social organisation, og i Santa Rosa National Park, Costa Rica, undergruppe størrelse kan variere fra 1 til 35 personer, men i gennemsnit omkring 5 personer findes rejser sammen. For denne population kan 50% af variansen i gennemsnitlig månedlig undergruppestørrelse forudsiges ud fra relativt rå målinger af størrelse, densitet og distribution af madplaster (Chapman 1990).

ud over de ovennævnte arter er anvendeligheden af den økologiske begrænsningsmodel generelt understøttet med forskning, der kommer fra en række arter (Snaith& Chapman 2005) og situationer (f.eks. 1993, og blandede arter foreninger, Chapman & Chapman 2000C). Vi mener, at tiden i betragtning af den omfattende støtte, som denne model har modtaget, er inde til, at det er rimeligt at foreslå, at arter typisk skal overholde forventningerne til modellen for økologiske begrænsninger. Dette betyder ikke, at alle arter vil tilpasse sig, og dermed bliver det en spændende tid for forskning på dette område, fordi efterforskere kan søge efter undtagelser fra modellen. Vi formoder, at arter, der ikke er i overensstemmelse, vil gøre det, fordi de har vedtaget sociale strategier, der strider mod de økologiske forventninger. Dette giver en” målestok ” for at evaluere den potentielle betydning af forskellige sociale strategier (dvs.jo længere man afviger fra forventningerne til den økologiske model, jo vigtigere er det sociale selektive pres).
lad os kort give to eksempler fra vores tidligere forskning. Kvindelige chimpanser var mere ensomme end kvindelige edderkopaber, alligevel har de begge lignende fission-fusion sociale organisationer. Yderligere, mens vi nøjagtigt kunne forudsige antallet af mænd og subadult chimpanser i en undergruppe baseret på økologiske forhold, var vi ikke i stand til at forudsige antallet af kvindelige chimpanser i disse undergrupper (Chapman et al. 1995). Selv når ressourcerne var ekstremt rigelige, og næsten alle mænd var i en stor undergruppe, kom kvinder sjældent ind i grupper, hvilket tyder på, at omkostningerne ved at være i en større undergruppe opvejer eventuelle fordele, såsom rovdyrundgåelse. Dette er uventet ud fra det perspektiv, at disse kvinders spædbørn ville være den alder/kønsklasse, der er mest truet af rovdyr. En testbar hypotese til at forklare disse observationer er, at koalitionernes art i disse to arter kan påvirke fordelene ved gruppemedlemskab. Bevis tyder på, at i modsætning til chimpanser danner edderkoppeabehunner koalitioner, der ofte fungerer for at give medlemmerne af koalitionen eksklusiv adgang til mad. Så edderkoppeaber, når de slutter sig til bestemte individer, kan øge deres adgang til mad ved at udelukke andre, men for chimpanse-hunner er dette ikke tilfældet (Chapman et al. 1995).
vi identificerede en anden interessant afvigelse, potentielt forårsaget af sociale faktorer, der påvirker omkostningerne og fordelene ved at være i en gruppe, og dette vedrører røde colobus og sort-hvide colobusaber (aka. i Kibale National Park, Uganda. Rød colobus danner store grupper med et gennemsnit på 65 individer (25-127 individer, Snaith et al. 2008), mens gueresas lever i små grupper med et gennemsnit på 6,5 individer (4-11 individer, Harris & Chapman 2007). På trods af denne forskel er der stor lighed i de planter, der spises af de to arter. Hvis økologiske forhold var ansvarlige for forskellen i gruppestørrelse mellem de to colobine-arter, ville man forvente, at deres kostvaner ville variere, og tætheden af madtræer ville være lavere i gueresas hjemområder, da de har den mindre gruppestørrelse. Imidlertid er deres kostvaner meget ens; kostoverlapningen mellem en rød colobus og en gueresa-gruppe, der havde sit hjemområde helt inden for hjemmeområdet for red colobus-gruppen, var 43.2%, Mens for de to tilstødende grupper af rød colobus, kost overlappet med kun 37,3% (Chapman et al. 2002). Det ser ud til, at Kvindelig gueresas reproduktive succes maksimeres i små og mellemstore grupper, enten fordi større grupper oplever højere overtagelser og barnemord eller mere fodringskonkurrence (se bevis for scramble konkurrence i folivorous Trachypithecus phayrei og Semnopithecus sp. , men mangel på bevis for, at gueresas nedbryder madplaster ). Vi har været vidne til en række mandlige overtagelser og spædbarnsdrab i gueresas (Onderdonk 2000)-se Teichroeb & Sicotte (2008) for en evaluering af teorien om barnemord i colobines. Hvad der ikke er klart er, hvordan risikoen for barnemord ville begrænse størrelsen af gueresa grupper, når de har tilsyneladende adgang til en høj tæthed af fødevarer, der bør tillade dannelsen af multi-mandlige grupper. I flere arter er større kvindelige grupper mere et mål for immigrerende mænd, og denne situation kan øge antallet af barnemord-for eksempel Semnopithecus sp. (Borries 1997), Theropithecus gelada (Dunbar 1984), Alouatta seniculus (Crockett & Janson 2000), Presbytis thomasi (Steenbeek & van Schaik 2001). Imidlertid forventes barnemord at være mindre hyppigt, hvis en gruppe indeholder flere reproduktivt aktive mænd, fordi mandlig indvandring og efterfølgende voldelig overtagelse og barnemord er mindre tilbøjelige til at forekomme, når den nytilkomne står over for flere mulige far til fremtidige spædbørn (Janson & van Schaik 2000). Gueresas ser ud til at have rigelige fødevareressourcer til at danne større grupper (Tombak et al. 2012), hvilket ville mindske risikoen for barnemord, men de udgør ikke store grupper. Det er muligt, at karakteren af mandlige-mandlige interaktioner i gueresas gør store grupper ustabile. Dette ser ud til at være tilfældet i ursine colobus (Colobus vellerosus), hvor multi-mandlige grupper tiltrækker mandlige indvandrere og har højere barnemordsrater end grupper med en stærk voksen mand i sin bedste alder (Teichroeb, Badescu, & Sicotte, upublicerede data; figur 3). Thus, even when ecological conditions would permit large groups to form, social conditions preclude their formation (Chapman & Pavelka 2005).

hvis den økologiske begrænsningsmodel skitserer pres, der generelt påvirker primatgruppestørrelse, og hvis afvigelser fra denne forventede gruppestørrelse skyldes socialt pres, vil det være en spændende tid på dette forskningsområde, fordi det rejser en række interessante spørgsmål. Eksempel: Hvad er karakteren af det sociale pres på gruppestørrelse? Hvorfor er disse sociale begrænsninger til stede i disse arter og ikke nært beslægtede? Og hvad er de evolutionære kræfter, der vælger for disse sociale pres? Gruppestørrelse er et kritisk element, der påvirker typen af social organisation arter vedtage, således at undersøge disse spørgsmål vil føre til spændende forbedringer af socioøkologiske modeller.
Leave a Reply